MINISTÈRE DE LA JEUNESSE, DE L'ÉDUCATION NATIONALE
ET DE LA
RECHERCHE
ÉCOLE PRATIQUE DES HAUTES ÉTUDES
Sciences de la Vie et de la Terre
THÈSE de
DOCTORAT
Mention :
SIEB
Spécialité
: Neurosciences Intégratives
présentée
publiquement par
Serge WUNSCH
Pour l'obtention du grade de
DOCTEUR de l'École Pratique des Hautes Études
Soutenue
le 21 novembre 2007
Rôle et importance
des processus
de renforcement dans l'apprentissage
du comportement de reproduction
chez l'homme
Evolution from mammalian heterosexual reproductive behavior to human erotic bisexuality
Abstract
What are the neurobiological dynamics of mammal and human sexuality? In rodents, mainly sexual hormones and pheromones, and secondarily sexual reflexes and reinforcement processes, would be the main innate factors at the origin of a genuine heterosexual reproductive behavior, the purpose of which is, at the favorable season, to achieve copulation permitting fecundation.
It would seem that during evolution, owing to the modifications of the brain of rodents into that of Man’s, the hormonal and pheromonal factors have become secondary, whereas the cognitive factors and the reinforcement processes (or reward systems) have become predominant. For these reasons, in Man, the behavior which leads to reproduction may be conceived not as an innate “reproductive behavior”, but as an acquired “erotic behavior” involving behavioral sequences of stimulation of the most erogenous body zones by a partner – no matter what his or her sex.
Reproductive behavior, sexual behavior, erotic behavior, bisexuality, heterosexuality, reinforcement, reward, erogenous zones, sexuality, Man.
Résumé
Quels sont les facteurs à l'origine des comportements humains ? Quels sont les processus neurobiologiques les plus primordiaux qui sous-tendent le développement et la dynamique de ces comportements ? Comment ces comportements émergent-ils de l'interaction entre les différents facteurs biologiques et environnementaux ?
Afin de répondre à ces questions capitales, nous avons réalisé une synthèse transdisciplinaire (neurosciences, éthologie, ethnologie et psychologie) de la littérature actuelle relative au comportement de reproduction. Ce comportement a été choisi car il est le comportement fondamental, absolument nécessaire à la survie de l'espèce, et, de ce fait, vraisemblablement soumis à une intense pression des mécanismes de la sélection naturelle. Pour cette raison, l'étude de ce comportement devrait permettre de mettre en évidence les principaux moyens biologiques qui ont été sélectionnés par l'évolution pour réaliser et contrôler les différents comportements fondamentaux de l'être humain.
À l'issue de l'analyse des données disponibles, il semblerait que les principaux facteurs biologiques innés (inhibition saisonnière par la mélatonine, hormones sexuelles, phéromones sexuelles, renforcements, lordose, érection, poussées pelviennes, réflexe éjaculatoire, libération de l'ovule lors du coït ...), à l'origine du comportement de reproduction des mammifères inférieurs, aient été modifiés au cours de l'évolution. Les facteurs hormonaux et phéromonaux seraient devenus marginaux tandis que les processus de renforcement et les facteurs émotionnels et cognitifs seraient devenus prépondérants. Apparemment, ce sont toujours les mêmes facteurs qui seraient à l'origine de la reproduction chez tous les mammifères, mais, comme l'importance relative de ces facteurs change au cours de l'évolution, la dynamique comportementale serait différente. En conséquence, chez l'Homme, l'innéité du comportement de reproduction aurait été perdue, et ce serait les processus de renforcement, associés aux zones érogènes, qui seraient devenus les principaux facteurs à l'origine de l'apprentissage d'un comportement permettant la reproduction. Ces données nous ont amenés à formuler l'hypothèse que c'est principalement l'intense érogénéité de la zone génitale (pénis/clitoris, vagin) qui provoquerait la découverte puis la répétition de différentes séquences motrices de stimulation des organes génitaux, dont la séquence du coït vaginal.
Pour des raisons éthiques, il n'a pas été effectué d'expérimentations neurobiologiques sur l'Homme. L'hypothèse a été testée au moyen d'un questionnaire, en utilisant la sensation consciente de plaisir érotique comme un indicateur de l'activité des processus de renforcement. Le questionnaire a été conçu de manière à pouvoir évaluer le rôle et l'importance du plaisir érotique – et donc indirectement des renforcements – dans l'apprentissage des activités de stimulation du corps et des zones érogènes, dont tout particulièrement l'activité du coït vaginal. L'enquête a été réalisée sur Internet au cours de l'année 2006 et 749 personnes ont complété le questionnaire.
L'analyse des résultats indique, principalement, que : 1) le plaisir érotique procuré par la stimulation par un partenaire des organes génitaux serait le plaisir le plus intense que l'organisme humain peut ressentir ; 2) les principales activités sexuelles s'organisent autour des zones les plus érogènes du corps, ce qui suggère que le plaisir érotique (et donc indirectement les renforcements) serait à l'origine de l'activité sexuelle ; 3) le coït vaginal, pourtant essentiel à la reproduction de l'espèce, n'est qu'une activité préférée parmi d'autres ; 4) les facteurs qui motivent les personnes a réaliser des activités sexuelles avec des partenaires seraient pour les deux tiers le plaisir somatosensoriel procuré par les différentes formes de stimulation du corps (contact corporel, étreintes, chatouilles, caresses sensuelles, stimulations érotiques ...), et pour un tiers des facteurs de type émotionnel (tendresse, complicité, curiosité ...) ; et, 5) il n'existe quasiment pas de différences significatives entre les réponses des femmes et des hommes, ce qui suggère l'existence d'une organisation neurobiologique des réactions sexuelles indépendante du genre de l'organisme.
En conclusion générale de l'analyse des résultats de l'enquête et de l'ensemble des données actuellement disponibles, il semblerait, dans l'espèce humaine, qu'il n'existerait plus de comportement de reproduction inné, mais, dû aux relations spécifiques et prépondérantes entre les processus de renforcement et les zones érogènes, il apparaîtrait un nouveau comportement dont le but est la stimulation du corps. Ce comportement, qui pourrait être qualifié d'érotique, induirait, indirectement, l'acquisition de la séquence cruciale du coït vaginal. Ainsi, la reproduction, pourtant fondamentale à la survie de l'espèce, ne serait paradoxalement chez l'Homme qu'une conséquence presque fortuite de la recherche des plaisirs physiques.
La généralisation des résultats de cette étude à l'ensemble des comportements humains suggère qu'il n'existerait pas d' "instincts" ou de "programmations" innés des comportements, mais plutôt tout un ensemble de processus neurobiologiques, innés mais élémentaires, à l'origine uniquement de tendances globales et approximatives, qui, au cours du développement et de l'interaction avec l'environnement, permettraient l'apprentissage par essais et erreurs de comportements relativement adaptés et appropriés à la survie de l'individu et de l'espèce.
Comportement de reproduction, comportement sexuel, comportement érotique, renforcement, récompense, zones érogènes, neurobiologie des comportements, instinct, inné / acquis, sexualité, plaisir, Homme.
1 – Théories relatives au comportement de reproduction
a – Description des comportements de reproduction
b – Préalable : définition des concepts clés
c – Théories actuelles et limites de ces théories
d – Le comportement de reproduction est-il inné ?
e – Apprentissage du comportement permettant la reproduction
2 – Présentation des hypothèses et du modèle comportemental
a – Présentation synoptique de l'hypothèse principale
b – Présentation détaillée du modèle comportemental
c – Comparaison entre les modèles de la reproduction
e – Possible extension du modèle et perspectives de recherche
f – Méthode de vérification de l'hypothèse principale
g – Transposition de l'hypothèse du niveau neurobiologique au niveau psychologique
a – Le plaisir érotique est-il un des plaisirs les plus intenses que l'organisme peut éprouver ?
d – Facteurs à l'origine des activités érotiques
a – Discussion relative aux limites de cette recherche
b – Discussion relative à la validité de l'expérimentation
c – Discussion relative aux résultats expérimentaux
d – Discussion relative à la validité de l'hypothèse et du modèle
e – Conclusion relative à la validité de l'hypothèse et du modèle
Annexe I : Proposition de définitions des concepts clés
Annexe III : Résultats et analyses de l'enquête par questionnaire
1 – Annexe du chapitre "Matériel & Méthode"
a – Critères de sélection des participants & Formations des groupes
b – Caractéristiques des participants à l'étude
b.1 – Profil des activités érotiques
2 – Annexe du chapitre "Résultats"
a – Le plaisir érotique est-il un des plaisirs les plus intenses que l'organisme peut éprouver ?
Annexe IV : Problèmes relatifs aux expériences éthiquement réalisables
"Connais-toi
toi-même"
Socrate
"La science vise à
l'amélioration du sort de l'Homme sur la terre."
Francis Bacon
We are recorders and
reporters of the facts. Not judges of the behavior we describe.
Alfred Kinsey
Remerciements
Je voudrais d'abord remercier toutes les personnes qui m'ont aidées, d'une manière ou d'une autre, à réaliser ces recherches.
Je remercie tout particulièrement le Dr. Marie-Christine Lombard pour avoir accepté, malgré les difficultés supplémentaires induites par ce travail pluridisciplinaire, de diriger ma thèse.
Je remercie également les Dr. Étienne Mullet, Jean-Pol Tassin et Olivier Rampin pour leurs critiques et conseils, ainsi que d'avoir accepté d'évaluer ce travail de recherche.
Je suis très reconnaissant au Dr. Philippe Brenot, qui a été, par ses conseils et son aide, à l'origine de cette thèse. Je le remercie également pour tous nos échanges constructifs.
J'exprime aussi une pensée amicale et chaleureuse au Dr. Christian Richard-Foy, pour ses apports cliniques et son soutient sans faille depuis le début de cette aventure intellectuelle.
Je remercie le Pr. Jacques Paty qui m'a donné l'opportunité d'une passation du questionnaire auprès de ses étudiants, ainsi que les Dr. Michèle Koleck et Romuald Nargeot pour leur aide concernant les analyses statistiques.
Je remercie également le Pr. Michel Le Moal qui m'a permis d'intégrer l'École Pratique des Hautes Études.
Enfin, je remercie ma famille qui m'a soutenu tout au long de ces années de recherches.
Liste des abréviations
A5: noradrenaline cells A5
Noyau noradrénergique A5
ACN: Anterior cortical nucleus (of amygdala)
Noyau cortical antérieur de l'amygdale
AOB: Accessory olfactory bulb
Bulbe olfactif accessoire
AON: Accessory olfactory nucleus
Noyau olfactif accessoire
Bar: Barrington's nucleus
Noyau de Barrington
BNST: Bed nucleus of the stria terminalis
Noyaux du lit de la strie terminale
CG: Central gray
Matière grise centrale
cl: colonne latérale (L6-S1)
CL: Central lateral nucleus of the thalamus
Noyau central latéral du thalamus
CM: Central medial nucleus of the thalamus
Noyau centromédian du thalamus
CPOF: Centroposterior orbitofrontal cortex
Cortex orbitofrontal centropostérieur (» aire 13)
DCN: Dorsal commissural nucleus
Noyau de la commissure dorsale
DLN: Dorsolateral nucleus
Noyau dorsolatéral
DM: Dorso median nucleus of the thalamus
Noyau dorsomédian du thalamus
DMvc: Caudal part of the medial dorsal nucleus of the thalamus
Partie caudale ventrale du noyau dorso-médian du thalamus
DPN: Dorsal penile nerve :
Nerf dorsal du pénis
DSTT: Dorsal spinothalamic tract
Faisceau spinothalamique dorsal
Ent: Entorhinal cortex
Cortex entorhinal
GL: Lateral geniculate nucleus of the thalamus
Corps géniculé latéral du thalamus
GM: Medial geniculate nucleus of the thalamus
Corps géniculé médial du thalamus
LD: Lateral dorsal nucleus of the thalamus
Noyau latéral dorsal du thalamus
LP: Lateral posterior nucleus of the thalamus
Noyau latéral postérieur du thalamus
LPOF: Lateroposterior orbitofrontal cortex
Cortex orbitofrontal latéropostérieur (» aire 12)
LTF: Lateral tegmental field (of mesencephalon)
Champ tegmental latéral
LVN: Lateral vestibular nucleus
Noyaux vestibulaires latéraux
MN: Medial nucleus (of amygdala)
Noyau médial de l'amygdale
MPO: Medial preoptic area
Aire préoptique médiale
MOB: Main olfactory area
Aire olfactive principale
NGc: Nucleus gigantocellularis (of reticular formation)
Noyau gigantocellulaire
nPGi: Nucleus paragigantocellularis
Noyau paragigantocellulaire
NPOm: Noyau préoptique médian
OT: Olfactory tubercule
Tubercule olfactif
Pf: parafascicular nuclei of the thalamus
Noyau parafasciculaire du thalamus
Pir: Piriform cortex
Cortex piriforme
PLCN: Posterolateral cortical nuclei (of amygdala)
Noyau cortical postérolatéral de l'amygdale
PMCN: Posteromedial cortical nucleus (of amygdala)
Noyau cortical postéromédial de l'amygdale
PP: Peripeduncular nucleus
Noyau péripédonculaire
PPRF: Paramedian pontine reticular formation
Formation réticulée pontique paramédiane
PVN: Paraventricular nucleus
Noyau paraventriculaire
RIMLF: Rostral interstitial nuclei of the median longi-tudinal fasciculus
Noyau interstitiel rostral du faisceau longitudinal médian
Rpa: Raphe pallidus nucleus
Noyau du raphé pallidus
Rmg: Raphe magnus nucleus
Noyau du raphé magnus
RS: Reticulo spinal
SI & SII: Somatosensory cortex I & II
Cortex somatosensoriel I & II
SNB: Spinal nucleus of the bulbocavernosus
Noyau spinal du bulbocaverneux
STT: Spinothalamic tract
Faisceau spinothalamique
TQA: Tubercules quadrijumeaux antérieurs
VMb: Basal part of the ventral medial nucleus of the thalamus
Région basale du noyau ventral médial du thalamus
VMpo: Posterior part of the ventral medial nucleus
Partie postérieure du noyau ventral médial du thalamus
VNO: Vomeronasal organ
Organe voméronasal
VPI: Ventral posterior inferior nucleus of the thalamus
Noyau ventral postéro inférieur du thalamus
VPL: Ventral posterior lateral nucleus of the thalamus
Noyau ventral postéro latéral du thalamus
VPLc: Caudal part of the ventral posterior lateral nucleus of the thalamus
Partie caudale du noyau ventral postéro latéral
VPM: Ventral posterior median nucleus of the thalamus
Noyau ventro-postéro-médian du thalamus
VSTT: Ventral spinothalamic tract
Faisceau spinothalamique ventral
Introduction
Le sujet de cette thèse s'inscrit dans le cadre général et fondamental de la recherche et de la compréhension des facteurs biologiques, environnementaux et culturels qui sont à l'origine des différents comportements humains.
Objectif et objet de la recherche
L'objectif principal de cette recherche transdisciplinaire (DELATTRE 1995) est d'identifier, chez l'Homme, les facteurs biologiques primordiaux qui sont à l'origine du comportement de reproduction. Ce comportement a été choisi car il est le comportement fondamental, absolument nécessaire à la survie de l'espèce, et, de ce fait, vraisemblablement soumis à une intense pression des mécanismes de la sélection naturelle. De plus, la reproduction chez les mammifères nécessite un minimum de coordination et de complémentarité entre les comportements de deux organismes de sexe opposé. Pour ces raisons, ce comportement semble être un objet d'étude privilégié permettant de mettre en évidence les principaux moyens biologiques qui ont été sélectionnés par l'évolution pour réaliser et contrôler les différents comportements fondamentaux de l'être humain.
Principal résultat
Les résultats de cette recherche suggèrent que l'anatomie et la physiologie de la reproduction seraient innées, tandis que le comportement de reproduction serait acquis.
Cette acquisition serait réalisée essentiellement grâce aux processus de renforcement, qui seraient à l'origine de l'apprentissage d'un comportement dont le but est la stimulation du corps, et non la reproduction. Le coït vaginal reproducteur serait ainsi une conséquence indirecte et presque fortuite de la recherche des plaisirs somatosensoriels.
Particularités de ce travail
Les deux principales difficultés de ce travail de synthèse transdisciplinaire (neurobiologie, éthologie, ethnologie et psychologie) sont l'ampleur du sujet étudié (la reproduction et la sexualité) et l'absence de recherche directe sur l'être humain dans ces domaines.
Élaborer et tester une hypothèse qui concerne un comportement dans sa globalité nécessitent, d'une part, de vérifier plusieurs hypothèses intermédiaires, et, d'autre part, de recueillir un grand nombre de données provenant de différents champs disciplinaires. L'analyse et la synthèse de ces nombreuses données et concepts pluridisciplinaires posent des problèmes spécifiques.
Par ailleurs, pour des raisons éthiques, il est impossible de tester des hypothèses neurobiologiques sur l'Homme. La vérification des hypothèses, qui ne peut donc être qu'indirecte, consiste à rechercher, de manière systématique et exhaustive, l'absence de réfutation et la maximalisation des corroborations.
Plan de l'ouvrage
Le premier chapitre, l'analyse de la littérature, est une analyse critique des différentes théories actuelles, explicatives du comportement de reproduction (voir page 3 ). L'objectif de ce chapitre est de mettre en évidence les limites de ces modèles et de présenter les données cruciales à l'origine de l'hypothèse de l'apprentissage du comportement permettant la reproduction.
Le second chapitre présente cette hypothèse de travail (voir page 50 ). L'objectif est de décrire l'hypothèse, et, afin de l'expliciter au mieux, de présenter également le nouveau modèle comportemental qu'elle implique. De plus, ce chapitre contient aussi des sous-chapitres nécessaires à la compréhension des particularités de cette recherche.
Le troisième chapitre, matériel & méthode, contient une description de la méthode et des moyens qui ont été utilisés pour tester et vérifier l'hypothèse principale (voir page 64 ). L'expérimentation a été réalisée au moyen d'un questionnaire évaluant l'importance des processus de renforcement dans la dynamique du comportement sexuel.
Les résultats de la partie expérimentale sont présentés dans le quatrième chapitre (voir page 68 ).
Le cinquième chapitre, discussion, concerne l'évaluation et l'argumentation des méthodes, des analyses et des résultats de cette étude (voir page 105 ). L'objectif est de préciser la validité et les limites des conclusions de cette recherche.
Les principaux résultats et analyses sont synthétisés dans la conclusion générale (voir page 147 ).
Enfin, les annexes contiennent différents documents complémentaires, apportant des explicitations et des précisions supplémentaires à certaines analyses ou résultats présentés dans les cinq principaux chapitres (voir page 169 et suivantes).
 |
1 – Théories relatives au comportement de reproduction
Les objectifs de ce chapitre sont : 1) de présenter les faits, c'est-à-dire les comportements qui permettent la reproduction ; puis 2) de présenter les théories, c'est-à-dire les structures d'idées qui permettent d'interpréter et d'expliquer ces comportements de reproduction ; puis 3) de mettre en exergue leurs limitations ; et enfin, 4) de présenter les principales données à l'origine de l'hypothèse de l'apprentissage du comportement permettant la reproduction.
a – Description des comportements de reproduction
La première étape de l'étude du comportement de reproduction est de décrire les séquences comportementales qui constituent ce comportement. Cette première étape, la description des faits, est distincte d'une seconde étape, l'élaboration d'une théorie, qui est l'explication de ces faits. La description des faits nécessite une grande rigueur afin de prendre en compte tous les faits significatifs existants dans les différents contextes et environnements. Pour l'Homme, en raison de la très grande influence du contexte culturel sur le comportement de reproduction, la description des faits tient compte de la diversité socioculturelle et est réalisée à partir des informations disponibles provenant de quelques centaines des 3.000 sociétés humaines connues (H.R.A.F. : Human Relations Area Files, banque mondiale de données ethnographiques, http://www.yale.edu/hraf/index.html).
Bien que l'étude du comportement de reproduction des animaux non humains ne soit pas l'objet de cette recherche, le comportement sexuel des rongeurs et des chimpanzés est présenté. Les données provenant de ces animaux sont les plus significatives pour cette étude, car les rongeurs sont la source de la majorité des données expérimentales et le chimpanzé pan paniscus (Bonobo) est l'espèce animale la plus proche de l'Homme.
Chez les rongeurs, le comportement de reproduction est relativement stéréotypé. La première phase du comportement de reproduction, appelée phase motivationnelle, permet un échange de stimulations adéquates entre les partenaires. Ces stimulations sont souvent nécessaires à la poursuite du comportement sexuel. En général, chez le rat, on observe les séquences suivantes : le rat mâle effectue une investigation olfactive, puis éventuellement gustative de la femelle, plus particulièrement de sa région périnéale. Cette investigation est d'autant plus longue que le rat est sexuellement inexpérimenté. Il se frotte parfois contre la femelle, et passe en dessous ou au dessus d'elle. Au cours de ces frottements, on observe fréquemment un comportement apparemment de marquage de la femelle par l'urine du mâle. Les deux partenaires peuvent émettre des vocalisations ultrasoniques. La rate œstrale répond aux stimulations du mâle par un comportement de saut et de fuite. En cas de défaillance du mâle, la femelle peut également prendre l'initiative en poussant les flancs de son partenaire. Puis, lors de la deuxième phase du comportement de reproduction, appelée phase consommatoire, le rat se positionne sur le dos et à l'arrière de la femelle. Durant la monte, le mâle palpe et étreint les flancs de la femelle avec ses pattes antérieures. La femelle est active et manifeste sa réceptivité par l'adoption d'une position de lordose et par l'orientation correcte de sa région périnéale, ce qui facilite l'intromission. Le mâle effectue des poussées pelvienne qui permettent l'intromission du pénis et l'éjaculation. Chez le mâle, l'éjaculation s'accompagne de contractions spasmodiques des muscles squelettiques. Après la copulation, le mâle procède généralement à une toilette de sa région génitale puis entre dans une période d'inactivité. L'éjaculation est suivie chez le mâle d'une période réfractaire caractérisée par une très faible réceptivité a tout type de stimuli et par un état de veille calme proche du sommeil (MEISEL & SACHS 1994).
Dans toutes les espèces sub-primates, on observe que le comportement de reproduction est principalement hétérosexuel et qu'il est saisonnier. Les femelles ne présentent pas de comportement sexuel avant la puberté, tandis que les mâles de beaucoup de ces espèces exécutent fréquemment des séquences incomplètes du comportement sexuel, longtemps avant leur maturité (FORD & BEACH 1965).
Chez le chimpanzé pan paniscus (Bonobo), en résumant l'essentiel, sa sexualité est bisexuelle (environ 1/3 d'activités homosexuelles et 2/3 hétérosexuelles, mais jamais d'homosexualité ou d'hétérosexualité exclusive), et on observe beaucoup d'activités non-reproductives : masturbation, utilisation d'objets, masturbation réciproque, baiser, activités oro-génitales, en groupe, etc. (ces activités représentent environ les 3/4 de toutes les activités sexuelles). Enfin, la sexualité est continue tout au long de l'année (DE WAAL 1992, 1996, 1990 ; HASHIMOTO 1997).
Chez l'Homme, le comportement sexuel est encore plus diversifié et on remarque des variations importantes en fonction des cultures. On observe des activités bisexuelles (caractérisée par des attitudes mâle et femelle, et/ou des partenaires mâle et femelle), et des activités hétérosexuelles ou homosexuelles, qui peuvent être exclusives ou non. On observe des activités autoérotiques, ou avec un ou plusieurs partenaires (de manière successive ou simultanée), comprenant des activités oro-orales, oro-génitales, génito-génitales ou anales, des caresses sensuelles, l'utilisation d'objet ou d'aliments, etc. (pour une description de ces activités, voir RÉMÈS 2004), et, lorsque le contexte culturel est favorable, on observe également la combinaison de nombreuses activités sexuelles avec d'autres activités hédoniques dans les orgies, ainsi que des activités sexuelles avec des animaux (FORD & BEACH 1965 ; ALLGEIER & ALLGEIER 1992 ; PARTRIDGE 2002). Les activité sexuelles humaines peuvent débuter dès l'âge de 4-5 ans, soit bien avant la puberté (voir les références dans la section "Remarques générales" ci-dessous). Enfin, les activités sexuelles peuvent être réalisées pour le plaisir, la procréation, la conformité aux normes sociales ou dans le cadre de rituels, de pratiques religieuses ou spirituelles. Il est difficile d'évaluer la part respective de chaque type d'activité, dans la mesure où l'influence culturelle est à l'origine de variations qui peuvent être majeures. Par exemple les activités homosexuelles sont très minoritaires dans les sociétés occidentales, mais représentent 100% des activités sexuelles avant le mariage dans certaines sociétés préindustrielles.
Pour ces raisons, dans cette étude, nous conserverons au terme « sexuel » sont sens général, et nous appellerons « comportements sexuels » l'ensemble de tous les comportements en relation avec les zones érogènes et la reproduction, « comportements de reproduction » les comportement sexuels dont le but est la copulation, « comportements érotiques » les comportement sexuels dont le but est la stimulation du corps et des zones érogènes – et non le coït vaginal –, et « sexualité » l'ensemble des aspects comportementaux, émotionnels et cognitifs liés aux comportements sexuels, qu'ils soient érotiques ou de reproduction.
Remarques générales
Les comportements sexuels décrits ci-dessus ne correspondent pas à une théorie, mais à des faits, qu'il sera absolument nécessaire de prendre en compte pour élaborer une théorie qui puisse les expliciter. Sous réserve que ces faits rigoureusement analysés correspondent à la réalité (car le recueil d'un fait résulte souvent d'une interprétation subjective, et parfois de biais moraux ou culturels (WALLEN & PARSONS 1997)), toutes les théories qui ne peuvent expliquer ces faits sont très probablement erronées.
Par ailleurs, on observe chez les primates que la fréquence des divers comportements sexuels n'est pas égale. Faut-il alors ne prendre en compte ou ne considérer comme significatifs que les comportements les plus fréquents, ou les plus culturellement valorisés ? En étudiant par exemple de manière détaillée le comportement sexuel qui semble le plus inapproprié à la reproduction, le plus "contre-nature", c'est-à-dire celui entre deux espèces distinctes, on s'aperçoit chez l'Homme que ce comportement dépend principalement du contexte. Bien que cette activité soit globalement peu pratiquée, il existe néanmoins quelques sociétés où l'activité sexuelle avec des animaux est culturellement acceptée. Mais le plus remarquable est que lorsque les êtres humains vivent avec les animaux, lorsqu'il existe une proximité physique et émotionnelle, et lorsqu'il n'existe pas d'interdits ou de stigmatisations culturelles, alors la probabilité d'observer ce comportement est très grande (KINSEY & al. 1948 ; FORD & BEACH 1965). Par ailleurs, les activités sexuelles entre animaux d'espèces différentes existent également, et elles sont d'autant plus fréquentes que la morphologie des organismes est similaire (âne/cheval, tigre/lion, etc.) (FORD & BEACH 1965). Cette remarque n'est pas une "éloge de la bestialité", mais un rappel de la nécessité à prendre en compte tous les faits observés, qu'ils soient fréquents ou marginaux, magnifiés ou dévalorisés, afin que la théorie élaborée puisse les expliciter et ainsi correspondre à la réalité.
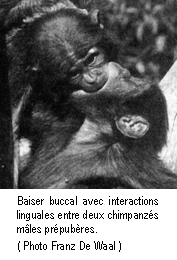 Enfin, dans l'objectif de comprendre
le développement du comportement sexuel et également d'évaluer les effets
supposés majeurs des hormones, des phéromones et de la puberté (cf. la
description des principales théories actuelles dans les sections suivantes), la
sexualité prépubère a été étudiée en détail chez les primates. En résumé, en
présentant l'essentiel, l'échographie montre que l'érection existe chez l'homme
bien avant la naissance (BROUSSIN & BRENOT 1996). Des études, menées en
particulier durant la période culturellement favorable des années 70,
confirment que l'organisme humain est capable de réactions et d'activités
érotiques dès le plus jeune âge (KINSEY & al. 1948 ; JOHNSTON
1973 ; CONSTANTINE & MARTINSON 1981 ; MARTINSON 1994). Par
ailleurs, les données ethnologiques indiquent que dans les sociétés où il
n'existe pas de restrictions culturelles à la sexualité des enfants
(Marquisiens (SUGGS 1966), Maori, Trobriandais (MALINOWSKI 1970), Pilagá (HENRY
& HENRY 1974), etc.), l'activité sexuelle débute vers 4-5 ans et les
enfants acquièrent le répertoire comportemental sexuel adulte, incluant le coït
vaginal, avant leur dixième année (FORD & BEACH 1965). De plus, on observe
des similitudes comportementales entre ces jeunes enfants et les jeunes
chimpanzés pan paniscus (Bonobo), tant dans le développement que dans
les activités du comportement sexuel. Les activités sexuelles des Bonobos,
espèce animale la plus proche de l'Homme, débutent vers l'âge d'un an, soit
bien avant la puberté, et ces activités prépubères représentent presque 1/4 de
la totalité des activités sexuelles (les activités des adolescents représentent
environ la moitié, et celle des adultes un peu plus du quart) (HASHIMOTO
1997 ; DE WAAL 1992, 1996, 1990). Toutes ces données suggèrent que
l'organisme des primates hominoïdes, bien avant les effets hormonaux et
phéromonaux de la puberté, est capable d'activités sexuelles dès les
premières années de la vie.
Enfin, dans l'objectif de comprendre
le développement du comportement sexuel et également d'évaluer les effets
supposés majeurs des hormones, des phéromones et de la puberté (cf. la
description des principales théories actuelles dans les sections suivantes), la
sexualité prépubère a été étudiée en détail chez les primates. En résumé, en
présentant l'essentiel, l'échographie montre que l'érection existe chez l'homme
bien avant la naissance (BROUSSIN & BRENOT 1996). Des études, menées en
particulier durant la période culturellement favorable des années 70,
confirment que l'organisme humain est capable de réactions et d'activités
érotiques dès le plus jeune âge (KINSEY & al. 1948 ; JOHNSTON
1973 ; CONSTANTINE & MARTINSON 1981 ; MARTINSON 1994). Par
ailleurs, les données ethnologiques indiquent que dans les sociétés où il
n'existe pas de restrictions culturelles à la sexualité des enfants
(Marquisiens (SUGGS 1966), Maori, Trobriandais (MALINOWSKI 1970), Pilagá (HENRY
& HENRY 1974), etc.), l'activité sexuelle débute vers 4-5 ans et les
enfants acquièrent le répertoire comportemental sexuel adulte, incluant le coït
vaginal, avant leur dixième année (FORD & BEACH 1965). De plus, on observe
des similitudes comportementales entre ces jeunes enfants et les jeunes
chimpanzés pan paniscus (Bonobo), tant dans le développement que dans
les activités du comportement sexuel. Les activités sexuelles des Bonobos,
espèce animale la plus proche de l'Homme, débutent vers l'âge d'un an, soit
bien avant la puberté, et ces activités prépubères représentent presque 1/4 de
la totalité des activités sexuelles (les activités des adolescents représentent
environ la moitié, et celle des adultes un peu plus du quart) (HASHIMOTO
1997 ; DE WAAL 1992, 1996, 1990). Toutes ces données suggèrent que
l'organisme des primates hominoïdes, bien avant les effets hormonaux et
phéromonaux de la puberté, est capable d'activités sexuelles dès les
premières années de la vie.
b – Préalable : définition des concepts clés
La réalisation de cette recherche concernant les facteurs primordiaux à l'origine du comportement de reproduction a nécessité de formuler des définitions opérationnelles des concepts d' "innéité", d' "instinct" et de "comportement". Ces définitions, utilisées dans cette étude, ont été élaborées à partir des connaissances anatomo-fonctionnelles et développementales des organismes biologiques.
La définition du concept d' "innéité" est la suivante : est "inné" tout élément (structurel ou fonctionnel) dont l'information développementale préexiste dans l'organisme et dont le développement est indépendant de l'influence des différents environnements (du chorion, de l'utérus, du milieu écologique, familial et culturel). Cet élément est toujours présent à la fin du développement, quelles que soient les caractéristiques de ces différents environnements, sauf dans les cas extrêmes où des caractéristiques particulières ont provoqué un développement pathologique (famine, maladies graves, …), qui dépasse les capacités adaptatives de l'organisme (physiologie du jeûne, système immunitaire, …).
La définition de l'instinct, au sens strict, est ainsi formulée : action ou réaction comportementale, innée, fixe, immédiatement fonctionnelle et adaptée sans expérience préalable et sans apprentissage, et provoquée par des stimuli internes ou externes spécifiques. Ces réactions sont déterminées par l'existence de structures neurales spécifiquement organisées (récepteurs spécifiques, et/ou synapses spécifiques, et/ou neuromédiateurs spécifiques, et/ou précablage spécifique...), dont la formation est indépendante des influences du milieu extérieur à l'organisme. L'exemple type d'un comportement sexuel instinctif est la lordose lombaire (voir PFAFF & al. (1994) pour sa description détaillée chez la rate).
La définition du concept de "comportement", valable pour tous les organismes vivants, est : ensemble des mouvements organisés pour agir à l'extérieur de l'organisme. Chez l'Homme, en tenant compte de son organisation anatomo-fonctionnelle, la définition spécifique devient : Ensemble des mouvements produits par l'activation du système musculaire squelettique, sous contrôle de l'activité neurale motrice pyramidale et extrapyramidale.
Les données et les analyses utilisées pour l'élaboration de ces définitions sont présentées à l'annexe I, page 170.
c – Théories actuelles et limites de ces théories
Le comportement de reproduction est un comportement fondamental pour la survie des espèces. Aucune espèce sexuée ne peut se perpétuer sans qu'il n'existe en sa structure des éléments biologiques particuliers lui permettant de réaliser un comportement destiné à sa reproduction.
Quels sont, chez l'Homme, ces facteurs biologiques fondamentaux ? Quel est l'état actuel des connaissances concernant ces facteurs ? Quelles théories permettent de donner un sens global à ces connaissances actuelles, permettant ainsi de comprendre et d'expliquer la sexualité humaine ?
Les principales théories actuelles, extraites de la littérature, sont analysées et présentée dans les paragraphes suivants.
Les premières théories modernes
La première théorie marquante a été élaborée par Richard von Krafft-Ebing (1840-1902), qui était considéré de son vivant comme un des plus grands psychiatres de son époque. Son principal ouvrage, Psychopathia Sexualis, publié pour la première fois en 1882, abondamment pourvu de cas cliniques exemplaires, a été constamment réédité jusqu'à aujourd'hui. Sa théorie, principalement axée sur les aspects pathologiques de la sexualité, était basée sur l'existence d'un instinct sexuel. Toutes les activités sexuelles qui ne permettaient pas la reproduction, c'est-à-dire qui s'écartaient du coït vaginal hétérosexuel, étaient considérées comme pathologiques. Néanmoins les bases de sa théorie relevaient du postulat, puisque l'étude et la justification de l'instinct sexuel se limitait à l'affirmation de son existence : "La perpétuité de l'espèce humaine n'est pas laissé au hasard, ni au caprice des individus : un instinct naturel la garantit, et il réclame impérieusement, irrésistiblement satisfaction." (Krafft-Ebing 1882, réédition 1999).
Sigmund Freud (1856-1939) a élaboré une théorie globale du psychisme humain, où la sexualité avait une place centrale. Sa théorie psycho-sexuelle supposait que la libido (l’énergie sexuelle) sous-tendait toute activité humaine. Son ouvrage Trois essais sur la théorie de la sexualité, publié en 1905, ainsi que le reste de son œuvre, furent accueillis avec indignation et dérision. Néanmoins, la psychanalyse devint finalement la théorie psychique la plus influente de l'époque, et aujourd’hui encore, elle influence un grand nombre de ceux qui s’intéressent à la sexologie. Néanmoins, là encore, les fondements de la théorie relèvent du postulat : la libido correspond en effet d'après S. Freud à l'énergie "de ces pulsions qui ont à faire avec tout ce que l'on peut comprendre sous le nom d'amour", et la pulsion correspond à des "poussées" psychiques d'origine biologique.
On remarque ainsi que les principales théories sexuelles du XIXe siècle sont basées sur l'existence quasi dogmatique d'une pulsion sexuelle ou d'un instinct sexuel, dont la preuve de leur existence est constitué uniquement par leur absolue nécessité à exister.
L'erreur de ce raisonnement est que si la reproduction est effectivement absolument nécessaire à la survie de l'espèce et qu'il doit absolument exister des éléments biologiques innés permettant d'aboutir à la fécondation, cela n'implique pas la nécessité de motivations innées (pulsion) ou de séquences motrices innées (instinct) spécifiques de l'hétérosexualité et du coït vaginal. L'essentiel est que la fécondation soit réalisée, peut importe les moyens et la manière qui permettent sa réalisation.
Théories récentes
Les diverses théories récentes ne sont pas présentées en détail dans cette étude, dans la mesure où les processus biologiques à l'origine du comportement de reproduction ne sont jamais décrits et restent spéculatifs. Le lecteur intéressé trouvera dans l'ouvrage "Sexualité humaine" (ALLGEIER & ALLGEIER 1992, p. 62-84) un exposé plus complet et les références des auteurs originaux.
En résumé, les théories évolutionnistes expliquent le comportement sexuel comme étant le résultat optimisé de la sélection naturelle. Les processus qui sous-tendent le comportement de reproduction ne sont pas décrits, mais ils sont supposés avoir été optimisés au cours des millions d'années de l'évolution.
Les théories sociobiologiques sont relativement similaires. Les comportements sexuels existent tels qu'ils sont aujourd'hui parce qu'ils ont dans un passé lointain donné à certains individus une valeur sélective supérieure.
Les théories issues des travaux de Pavlov et de Skinner mettent l'accent sur l'influence des conditionnements et des récompenses dans le développement du comportement sexuel. Néanmoins, l'origine des stimuli inconditionnés qui permettent ces apprentissages n'est pas explicité.
En synthèse, on observe dans toutes ces théories de la sexualité humaine qu'elles sont quasiment toutes basées sur la fonction fondamentale de reproduction. Il est généralement admis, en raison de l'importance cruciale de la reproduction pour la survie de l'espèce, que les organismes sexués doivent certainement comporter des éléments biologiques spécifiquement prévus et organisés pour contrôler un comportement permettant la fécondation. L'évolution aurait sélectionné une organisation innée, adaptée et optimisée, afin que les organismes se reproduisent. Cette organisation "instinctuelle" – qui n'est jamais décrite – aurait comme principale et unique finalité la reproduction et serait à l'origine de séquences comportementales hétérosexuelles permettant le coït vaginal entre des partenaires fécondables. Les apprentissages joueraient un rôle important, mais concerneraient plutôt des caractéristiques secondaires. L'essentiel, c'est-à-dire la réalisation du coït vaginal hétérosexuel, serait inné. Même si actuellement le concept d' "instinct" n'est plus très usité, il reste néanmoins, en filigrane, la référence à partir de laquelle sont basés la plupart des théories actuelles.
Théories neurobiologiques
Le développement récent des neurosciences a permis l'exploration anatomique et fonctionnelle du système nerveux, ainsi que des fondements neurobiologiques des comportements. Néanmoins, malgré de nombreuses recherches concernant le comportement de reproduction, il n'existe pas de modèle neurobiologique global.
C'est ce qu'on observe par exemple dans les articles de synthèse publiés dans Annual review in sex research, le seul journal scientifique spécialisé dans les reviews en sexualité. Une analyse des publications de cette revue montre qu'aucun modèle global, ni biologique ni psychologique ou ni pluridisciplinaire, n'est publié.
Il apparaît que les différents chercheurs et laboratoires sont spécialisés dans un certain type d'étude (la différenciation sexuelle, les hormones, les phéromones ou les processus de renforcement, sur les rongeurs, les poissons ou les oiseaux, etc.), et les différents modèles publiés, partiels, reflètent bien souvent la spécialisation de leurs auteurs.
Par exemple, Thomas Insel est spécialisé dans l'étude de la formation des liens d'attachement entre les partenaires sexuels (INSEL 2003, 2001, 1997, 1995, 1992, 1991), Anders Ågmo est plutôt spécialisé dans l'étude des renforcements sexuels (AGMO 2003, 2002, 1999, 1995, 1993, 1990), Michael Meredith étudie principalement les phéromones (MEREDITH 2001, 1994, 1991), etc.
Dans un dossier récent de Nature neuroscience concernant la sexualité, le résumé introductif de Brian Fiske (2004) est représentatif des théories neurobiologiques actuellement dominantes, celles centrées sur les hormones sexuelles. Ces hormones seraient à l'origine de la formation au cours du développement d'un circuit (partiellement hypothétique) responsable de la motivation et du contrôle du comportement sexuel. À la puberté, la testostérone activerait ce circuit, qui contrôlerait, d'une part, la motivation à l'origine du rapprochement des mâles et des femelles, puis, d'autre part, le déroulement des séquences aboutissant au coït vaginal. Néanmoins, bien que ce dossier soit clairement destiné à apporter des informations concernant la sexualité humaine, l'essentiel des données concernent la reproduction, les hormones sexuelles et les relations hétérosexuelles chez l'animal. De plus, même les données animales connues relatives aux phéromones, aux renforcements, aux conditionnements et aux apprentissages ne sont ni analysées ni même citées afin de relativiser la portée des conclusions.
Dans leur ouvrage Psychologie biologique, qui est à la fois un ouvrage de synthèse pluridisciplinaire et une méthode d'étude et d'explication des comportements à partir de la biologie, Rozensweig & al. (2002) présentent la sexualité d'une manière qui est typique des ouvrages actuels de neurosciences. La première page (p. 363) est une introduction centrée sur la sexualité humaine, puis quatre pages (p. 366-369) concernent la copulation chez le rat, suivie de trois pages (p. 370-372) traitant essentiellement de l'aspect physiologique de la reproduction humaine, puis une page (p. 373) sur les phéromones, et enfin vingt-quatre pages (p. 374-397) concernent les aspects phylogénétiques et ontogénétique de la sexualité, avec une mise en exergue des effets des hormones sexuelles. Si pour le rat un modèle explicatif du comportement de reproduction est présenté (p. 369), pour l'Homme il est juste présenté des données cliniques et expérimentales indiquant dans certains cas particuliers des effets hormonaux et phéromonaux. Aucun modèle explicatif n'est présenté, la diversité du comportement sexuel humain est juste citée, les renforcements, associations et conditionnements ne sont pas abordés, et l'influence culturelle à peine évoquée. On observe que les auteurs ne présentent que les connaissances bien établies en biologie et éludent tous les aspects qui pourraient être considérés comme spéculatifs ainsi que tous les aspects de la sexualité qui n'ont pas un lien direct avec la reproduction. Les connaissances présentées sont valides et fiables, mais partielles. Les facteurs et la dynamique de la reproduction humaine ne sont que suggérés. L'impression qui se dégage après la lecture de ce chapitre est que les processus fondamentaux permettant le comportement de reproduction chez l'Homme sont vraisemblablement très similaires à ceux des rongeurs, avec certainement un effet prépondérant des hormones sexuelles. Quand aux aspects non reproductifs de la sexualité, qui sont à peine évoqués, on ne connaît ni les facteurs à leur origine, ni leurs relations avec les facteurs de la reproduction.
Néanmoins, après la synthèse des différents articles et ouvrages concernant la reproduction, il apparaît en résumé le modèle suivant du comportement de reproduction chez les mammifères : les hormones prénatales, et principalement la testostérone chez l'homme, seraient à l'origine du développement d'un circuit neural spécifique au comportement sexuel (phase d'organisation). Ce circuit, qui deviendrait mature entre la période de la puberté et l'âge adulte, serait responsable de la motivation sexuelle à rechercher un partenaire sexuel approprié et contrôlerait les séquences motrices conduisant au coït vaginal. À la puberté, l'augmentation importante du niveau des hormones sexuelles activerait ce circuit neural spécifique du comportement de reproduction (phase d'activation). Les phéromones, sous contrôle hormonal, permettraient la reconnaissance du partenaire approprié et l'initiation du comportement reproducteur. Les réflexes sexuels (lubrification vaginale, érection, poussées pelviennes, éjaculation, etc.) permettraient la réalisation des différentes séquences successives aboutissant à la transmission des gamètes et à la fécondation. Enfin, les renforcements et les conditionnements, l'expérience individuelle, des facteurs psychologiques et culturels moduleraient l'expression du comportement sexuel, mais celui-ci serait biologiquement déterminé pour l'hétérosexualité et le coït vaginal.
Insuffisances des théories actuelles
Mais ce modèle neurobiologique actuel, axé sur le coït hétérosexuel instinctuel et basé sur l'expérimentation animale, présente de nombreuses insuffisances et beaucoup de questions importantes, voire cruciales, restent sans réponses chez l'Homme :
– Quel est précisément le circuit neural responsable du comportement sexuel (localisation, constitution, dynamique fonctionnelle, ...) ?
– Quels sont les principaux processus neurobiologiques à l'œuvre ?
– Où et comment agit la testostérone, afin de réaliser le coït vaginal fécondant ?
– Quelle est la fonction et l'importance des processus de renforcement ?
– Quelle est la fonction du plaisir ?
– Quelle est l'importance de l'apprentissage, et quels aspects de la reproduction seraient acquis ?
– Comment expliquer les variations du comportement sexuel entre les espèces ?
– Comment expliquer les activités sexuelles non reproductrices :
– La masturbation.
– Le baiser.
– Les caresses sensuelles.
– Les activités oro-génitales (fellation, cunnilingus ...).
– Les activités bi- ou homo-sexuelles.
– Les activités sexuelles prépubères (qui représentent chez les chimpanzés pan paniscus presque 1/4 des activités sexuelles).
– Les activités sexuelles après la ménopause ou l'andropause.
– L'utilisation d'objets.
– Les activités sexuelles entre espèces (en particulier entre primates).
– Comment expliquer les activités sexuelles potentiellement reproductrices, mais atypiques, telle la sexualité de groupe ?
– Qu'est-ce que le phénomène appelé "amour" ? Quelle est sa fonction et son importance dans la sexualité ?
– Comment expliquer l'addiction sexuelle ?
La quantité et l'importance de ces insuffisances, ainsi que la nécessité d'adjoindre de nombreuses hypothèses complémentaires ad hoc afin d'expliciter ces insuffisances (indifférenciation de l'instinct avant la maturation, inversion de l'instinct, homosexualité latente ou contextuelle, plaisir sexuel, activités préliminaires d'amorçage, pathologies, perversions, etc.), mettent en question, au moins chez l'Homme, la validité des théories neurobiologiques actuelles.
Origine des insuffisances
Ces insuffisances des modèles théoriques contemporains découleraient de la conjonction de plusieurs facteurs :
– L'existence en Occident de valeurs et de représentations culturelles hétérocentristes.
– La dominance en sexologie d'un paradigme homosexuel/hétérosexuel qui rend difficile, voire inconcevable, l'existence d'autres formes de sexualité, telle par exemple la bisexualité (RODRIGUEZ-RUST 2003), ou d'autres conceptions théoriques radicalement différentes.
– L'utilisation de données expérimentales provenant essentiellement d'animaux ayant un comportement de reproduction très différent de l'Homme (rongeurs, oiseaux et poissons).
– La non prise en compte de toutes les activités sexuelles humaines qui ne permettent pas la reproduction. Ces activités sont généralement considérées comme relevant de contextes particuliers ou comme étant pathologiques.
– L'absence de recherches fondamentales et transculturelles de la reproduction et de la sexualité humaine.
d – Le comportement de reproduction est-il inné ?
Par ailleurs – et surtout – il semble que l'aspect essentiel de la plupart des théories actuelles, c'est-à-dire l'innéité du coït vaginal, soit réfuté tant chez l'Homme que chez les autres primates.
En effet, on observe que lorsque les mammifères non humains, et tout particulièrement les primates, sont mis dans des conditions où ils ne peuvent apprendre aucun élément du comportement de reproduction, de manière systématique le mâle est incapable de coïter (GRUENDEL & ARNOLD 1969 ; MISSAKIAN 1969 ; TURNER & al. 1969 ; WARD 1992 ; FORD & BEACH 1965). Cette incapacité du mâle à pratiquer le coït en dehors de toute expérience préalable a été vérifiée dans plusieurs espèces (cochon d'Inde, rat, chat, chien, macaque rhésus, chimpanzé), et elle est systématique chez tous les primates. En variant les conditions expérimentales, il est apparut que c'est vraisemblablement la privation du contact physique, et non de la vue, des sons ou des odeurs des congénères, qui est le facteur critique à l'origine du déficit coïtal (WARD 1992 ; GRUENDEL & ARNOLD 1969). On peut ainsi observer des mâles sexuellement naïfs manifester de nombreux comportements spécifiques de la reproduction (excitation, érection, contacts avec le partenaire ...) mais ils ne parviennent pas à copuler. Un des principaux problèmes, bien qu'il existe également des problèmes connexes de socialisation et de peur des autres congénères (GOLDFOOT 1977), pourrait être lié à un déficit de construction du "schéma corporel" et se caractérise par une incapacité à positionner correctement le corps afin de réussir l'intromission (HARD & LARSSON 1971).
De plus, ces déficits comportementaux sont corrélés avec un moindre développement de l'amygdale médiale et du noyau préoptique médian (NPOm). Ces données montrent que le développement de certaines régions sexuellement dimorphes, sensibles aux hormones sexuelles et supposées jouer un rôle critique dans le comportement de reproduction et la copulation, n'est pas inné et dépend de l'expérience sociale (COOKE & al. 2000).
En conclusion, toutes ces données suggèrent que chez les mammifères non humains, et systématiquement chez les primates, au minimum, le mâle doit apprendre la séquence la plus cruciale, celle du coït vaginal.
Par extrapolation, l'Homme n'ayant que quelques pour cent de différence génétique avec les autres primates, il est quasiment impossible, en fonction des connaissances actuelles, de concevoir comment des mutations sur un nombre restreint de gènes peuvent coder ex nihilo une séquence comportementale aussi spécifique que le coït vaginal. Ces données animales corroborent plutôt l'hypothèse de la non innéité du coït vaginal chez l'Homme.
Vérification de l'innéité : Recherche systématique d'éléments innés
Les données déterminantes présentées ci-dessus montrent que chez les primates la séquence cruciale du coït vaginal n'est pas innée chez le mâle. Par ailleurs, il est généralement admis que déjà chez l'animal les apprentissages ont une grande importance (PFAUS & al. 2001), que l'importance des hormones et des phéromones décroît avec le développement cortical (BUVAT 1996 ; FOIDART & al. 1994), et qu'une grande partie des comportements sexuels humains serait appris (ALLGEIER & ALLGEIER 1992). Toutes ces données suggèrent que chez l'Homme, l'essentiel, et peut être la quasi totalité du comportement de reproduction ne serait pas inné mais acquis.
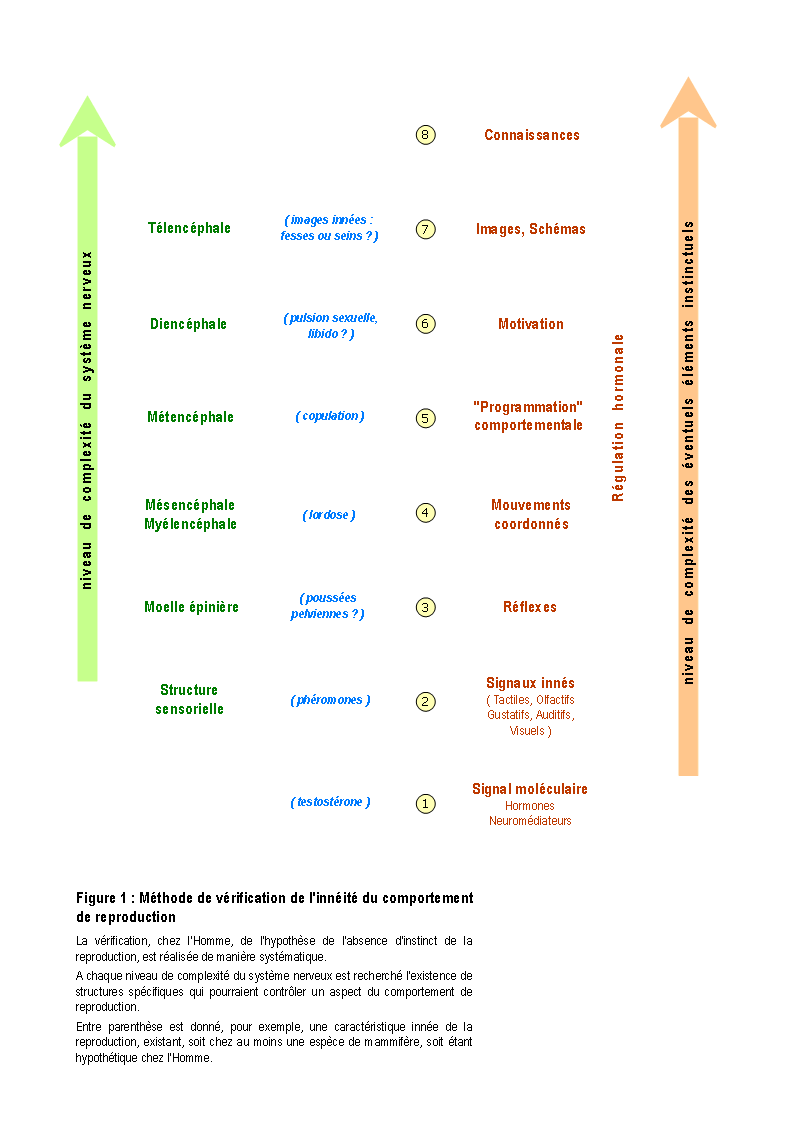
Afin de vérifier cette possibilité avec un maximum de validité, une analyse systématique et méthodique de la littérature a été réalisée pour savoir s'il existe des caractéristiques innées du comportement de reproduction. S'il en existe, les théories actuelles sont partiellement valides. S'il n'en existe pas, la majorité des théories actuelles sont réfutées et il serait alors vraisemblable que le comportement de reproduction humain soit entièrement acquis.
Dans les sections suivantes, la présence ou l'absence de différents processus innés, intervenant dans le comportement de reproduction et la séquence cruciale du coït vaginal, est recherchée de manière systématique dans l'ensemble du système nerveux (cf. également la section "Transcription biologique de l'instinct", en annexe, p. 172 et suivantes). Toutes les structures et les processus neuraux, des plus simples aux plus complexes, sont successivement explorés : systèmes neurohormonaux, systèmes sensoriels, système moteur, arcs réflexes, processus émotionnels et processus cognitifs (voire l'illustration de cette méthode dans la Figure 1, ci-contre).
Figure 1
a – Hormones et neuromédiateurs
Au niveau moléculaire, existe-t-il des molécules, des neuromédiateurs ou des hormones qui ont un effet inné sur le comportement de reproduction ? ( Figure 1, niveau [1] )
Introduction : molécules & mécanismes moléculaires
Dans un premier temps, une analyse générale des connaissances actuelles concernant les molécules agissant dans le système nerveux permet-elle d'identifier des molécules ayant des "propriétés sexuelles innées" ? Les résultats de cette synthèse relative à la communication cellulaire, qui n'est pas l'objet de ce travail de recherche, est très brièvement présenté dans ce paragraphe.
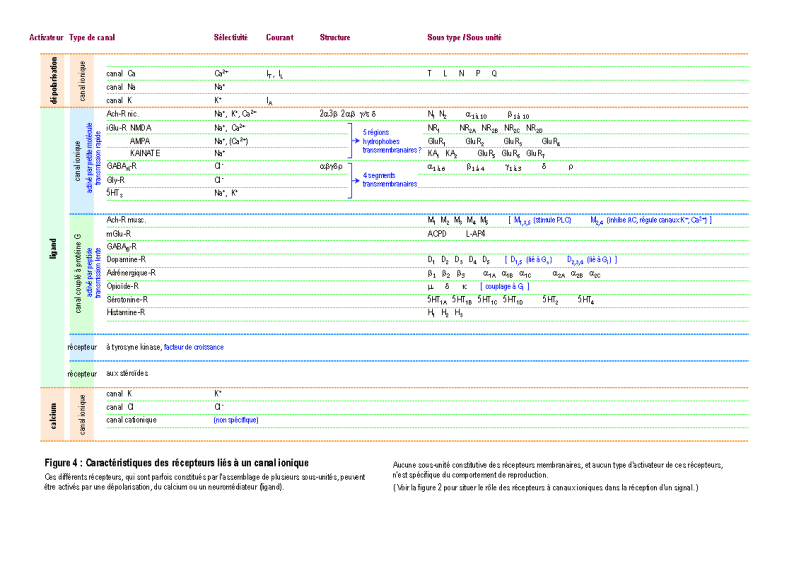
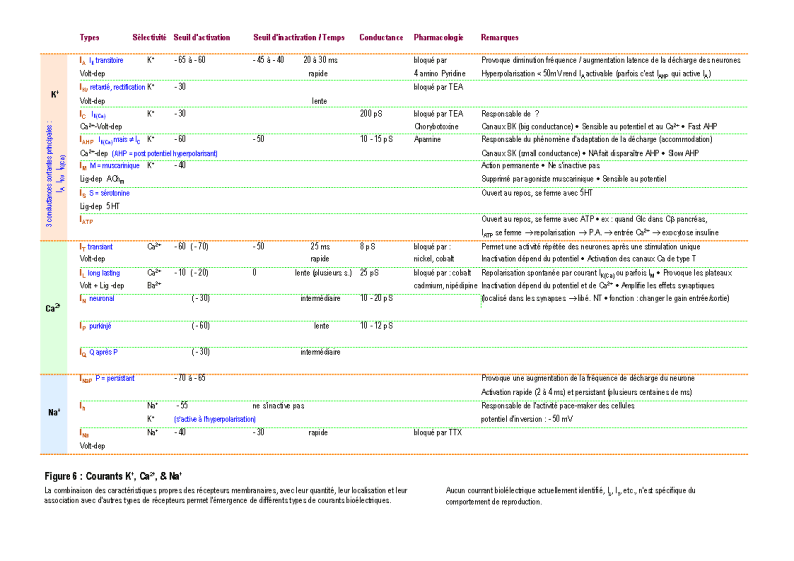
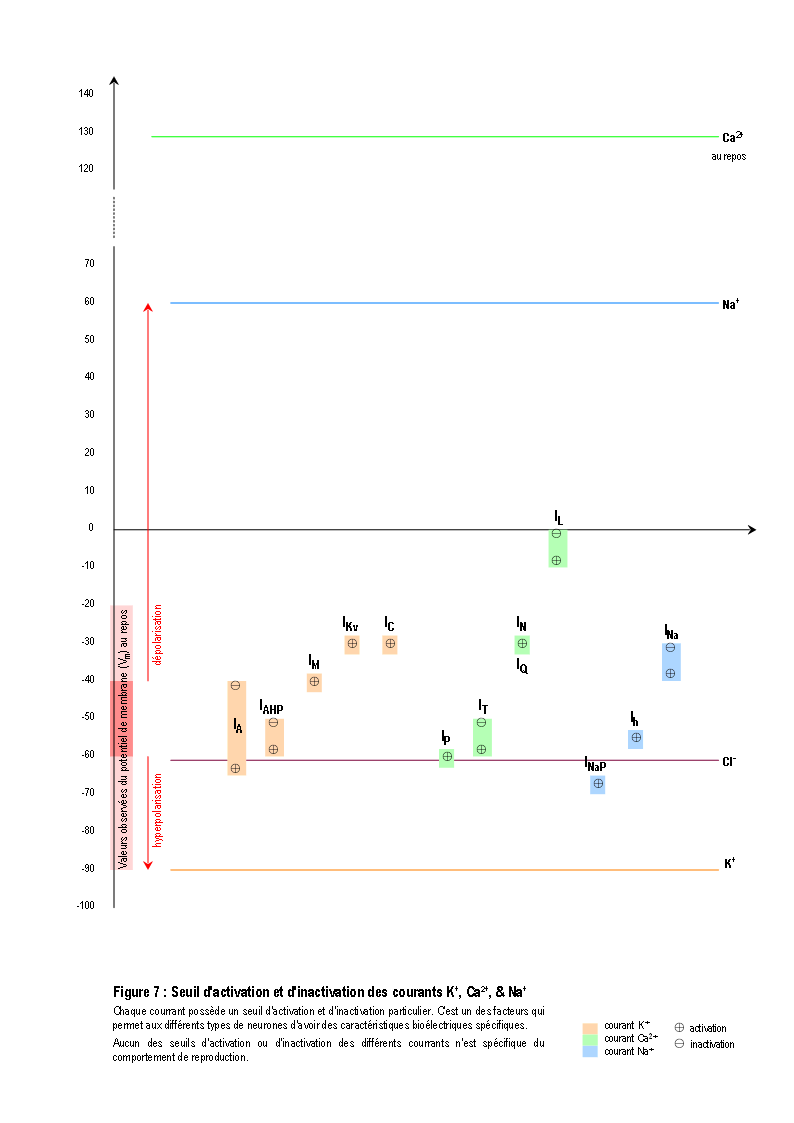
Une molécule, neuromédiateur ou hormone, peut avoir une fonction de transmission d'un signal en se fixant sur un récepteur membranaire ou intracellulaire d'une cellule (Figure 2). On connaît actuellement plus d'une centaine de molécules distinctes les unes des autres qui peuvent chacune véhiculer un signal particulier (voir la Figure 3 pour une présentation résumée des différents neuromédiateurs). La nature et l'effet du signal dépendent également de la structure des récepteurs, qui sont souvent constitués par un assemblage de différentes sous-unités (voir la Figure 4 pour une présentation résumée des différents types de récepteurs et de sous-unités). L'activation des récepteurs membranaires permet le passage d'ions ou la libération de seconds messagers (Figure 5), qui déclenchent des réactions cellulaires. La présence dans un neurone de différents récepteurs membranaires peut permettre de générer différents types de courant bioélectrique (Figure 6 et Figure 7). D'autres molécules, telles les hormones stéroïdes, pénètrent dans le neurone et se lient à des récepteurs nucléaires intracellulaires. Elles activent la transcription de certains gènes, induisant ainsi des activités métaboliques (en général la synthèse de protéines), qui modifient les propriétés et les activités des neurones (modifications électrophysiologiques, apoptose, remaniement du cytosquelette ...). Finalement, tous ces effets modifient soit l'activité métabolique du neurone, soit modifient l'architecture du neurone ou de celles de ses connexions, ou soit modifient ses propriétés bioélectriques. (pour des explications détaillées, voir : DARNELL & al. 1993 ; HAMMOND & TRITSCH 1990 ; KANDEL & SCHWARTZ 1995 ; RAWN 1990 ; SARGENT 1994 ; VAN OOYEN & VAN PELT 1994 ; WILSON 1999).
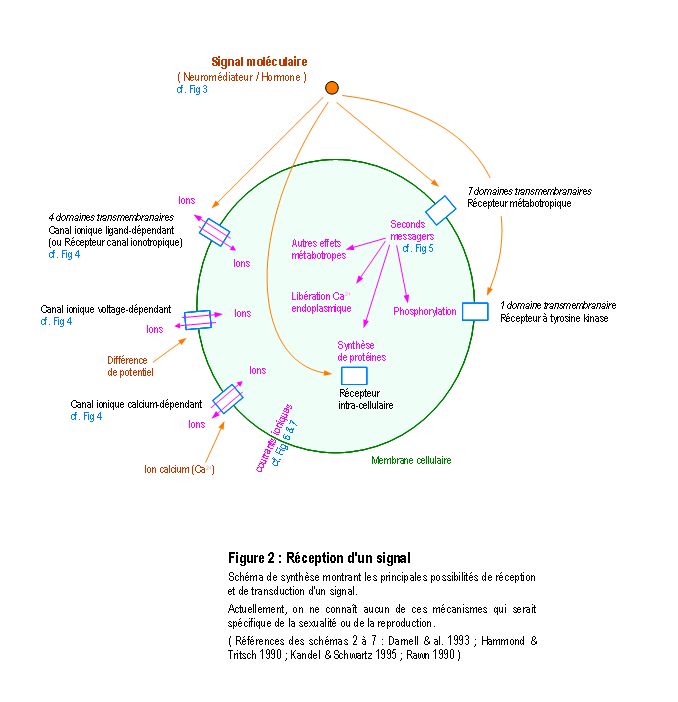

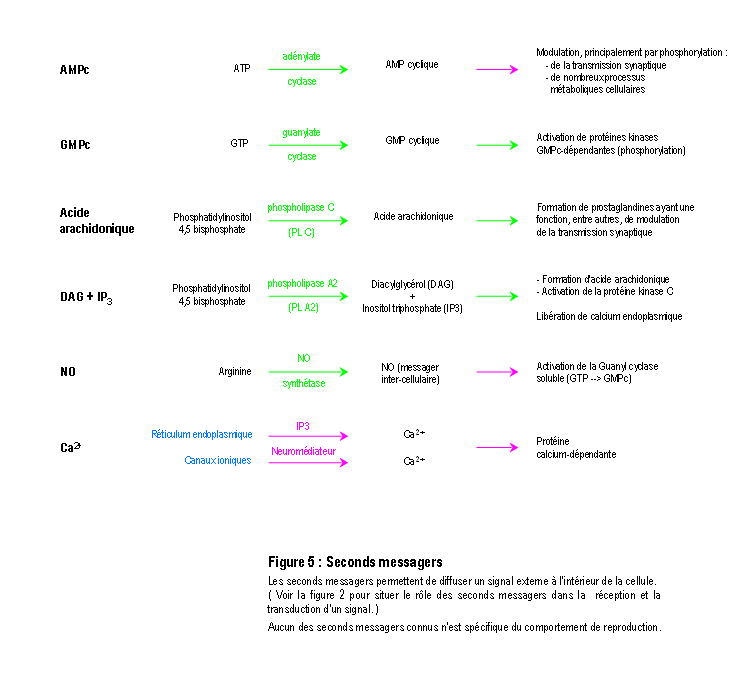
La compréhension détaillée de ces mécanismes moléculaires et cellulaires, où l'on n'observe aucune activité directement lié avec la reproduction, permet d'affirmer qu'en l'état actuel des connaissances il ne peut pas exister au niveau moléculaire et cellulaire de caractéristiques qui rendent une molécule spécifique et exclusive de la reproduction et qu'également aucun mécanisme moléculaire ou cellulaire n'est spécifique ou exclusif de la reproduction.
Dans un second temps, la compréhension de ces mécanismes d'action de la communication cellulaire permet de comprendre que les effets dans le système nerveux d'une hormone ou d'un neuromédiateur dépendent en fait essentiellement des caractéristiques structuro-fonctionnelles du neurone récepteur et du réseau créé par l'architecture de ses connexions : l'action sur des neurones sensoriels et moteurs produira, respectivement, des effets sensoriels dans le premier cas et moteurs dans le second cas. Et un effet moteur ne pourra être considéré comme inné et spécifique du comportement de reproduction que si l'organisation au sein de cette structure de la réception, du traitement et de la transmission ultérieure du signal sont également innés et spécifiquement prévus pour exécuter des séquences motrices aboutissant au coït vaginal.
En conclusion, on observe qu'il existe des molécules qui, en fonction de leurs cibles neurales, ont des effets qui peuvent être en relation avec la reproduction, mais qu'il n'existe ni de mécanismes moléculaires ni d'hormones ou de neuromédiateurs possédant des caractéristiques intrinsèques qui les rendent spécifiques uniquement du comportement de reproduction. S'il existe un instinct de la reproduction, il doit être recherché à un autre niveau d'organisation.
Remarque : les hormones dites "sexuelles"
Ces mécanismes neurobiologiques permettent d'expliquer les raisons pour lesquelles des hormones dites "sexuelles", telle la testostérone, qui ont des effets important sur la sexuation et le comportement de reproduction (MEISEL & SACHS 1994), ont également des effets sur les processus sensoriels (GANDELMAN 1983), les émotions (BEATTY 1992), et l'agression (KARLI 1987 ; GARIEPY & al. 1996).
Bien que la majorité des données expérimentales proviennent des rongeurs et que les données humaines sont rares, ces hormones dites "sexuelles" sont actuellement considérées par la plupart des auteurs comme étant chez l'Homme le facteur majeur à l'origine d'un comportement inné de la reproduction.
Mais en tenant compte des dernières données pluridisciplinaires, en particulier celles provenant des primates hominoïdes, cette conclusion est-elle toujours valide ? Quels sont chez l'Homme les sites d'action et les effets de ces hormones sexuelles sur le comportement de reproduction, et, surtout, quelle est leur importance relative comparée à tous les autres facteurs impliqués dans ce comportement ?
Chez l'Homme, on observe que les hormones sexuelles ont différents effets spécifiques en relation avec la sexualité :
– Différenciation sexuelle en organisme mâle et femelle (sexuation).
– Différenciation sexuelle des sites impliqués dans la morphologie sexuelle.
– Effets physiologiques (développement des gamètes, cycle menstruel ...).
– Différenciation sexuelle des sites de production, de réception et de traitement des phéromones.
– Différenciation sexuelle des sites impliqués dans des séquences motrices liées à la reproduction.
– Augmentation de la motivation sexuelle.
Les trois derniers effets pourraient être considérés comme étant des effets innés contrôlant le comportement de reproduction. Ces effets sont discutés dans les sections appropriées (réflexes sexuels, olfaction, motivation ...) des pages suivantes, en fonction de la région et du niveau d'organisation du système nerveux où agissent ces hormones dites "sexuelles".
b – Canaux sensoriels
La réalisation du coït vaginal nécessite obligatoirement un échange de signaux entre les partenaires. Il est donc possible de vérifier s'il existe des structures sensorielles qui permettent la réception et le décodage de signaux innés, spécifiques à la copulation ( Figure 1, niveau [2], page 14 ).
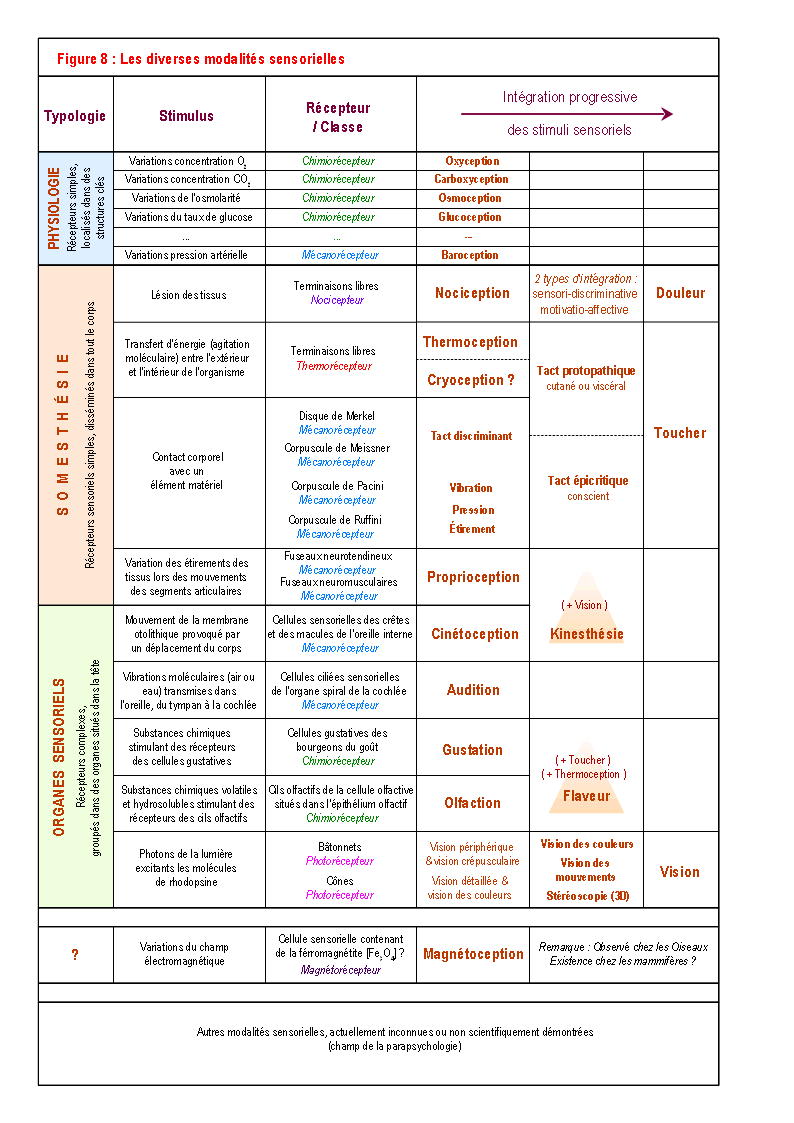
Dans un premier temps, une analyse générale des connaissances actuelles concernant les systèmes sensoriels permet-elle d'identifier des canaux de communication susceptibles de transmettre des signaux sexuels innés ? Les différents récepteurs, organes et systèmes sensoriels connus sont présentés de manière synoptique dans la Figure 8 (pour des explications détaillées, voir : ROSENSWEIG 1991 ; WILLIS & COGGESHALL 1991 ; BESSON & al. 1994 ; HOLLEY 1996). En fonction des données actuelles, les chimiorécepteurs et les mécanorécepteurs qui détectent certains signaux internes (oxygène, dioxyde de carbone, glucose ...), permettant ainsi le maintien de l'homéostasie, ne semblent pas pouvoir être impliqué dans la détection, même indirecte, de signaux innés du comportement de reproduction. Ils ne sont donc pas pris en compte dans cette étude. Par contre, les structures sensorielles de la somesthésie et des organes sensoriels peuvent percevoir des stimuli externes, et, de plus, soit possèdent des récepteurs spécifiques (par exemple spécifique à une phéromone sexuelle) ou soit rendent possible l'analyse au niveau cortical d'un signal complexe (forme visuelle par exemple) permettant de faire correspondre les caractéristiques prototypiques de ce signal à une éventuelle référence interne innée. Pour ces raisons, l'olfaction, la vision, l'audition, le toucher et le goût sont des canaux sensoriels qui peuvent recevoir un signal inné, et sont donc étudiés en détail dans les sections suivantes.
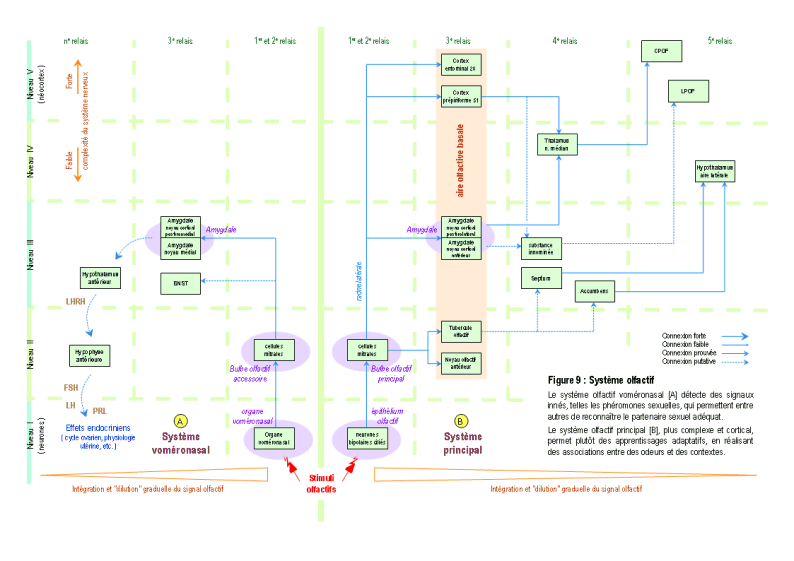
b.1 – Olfaction
Introduction : processus olfactifs
Il existe chez l'Homme deux systèmes olfactifs, principal et voméronasal (STENSAAS 1991), susceptibles de détecter des phéromones (MEREDITH 2001) ( Figure 9, partie [A] et [B] ) (pour des explications détaillées, voir : FIRESTEIN 2001 ; NIEUWENHUYS & al. 1988 ; BOSSY 1990 ; SHIPLEY & al. 1996 ; SIGNORET 1996 ; HOLLEY 1996 ; SCHAAL 1996). Les récepteurs olfactifs (cils olfactifs de la cellule neurosensorielle de l'épithélium olfactif, et les microvillosités des cellules sensorielles de l'organe voméronasal) sont les seuls récepteurs de l'organisme mammalien qui peuvent détecter à distance des signaux spécifiques. En effet, une molécule olfactive peut provenir d'un autre organisme et peut avoir des caractéristiques physiques et chimiques particulières et bien distinctes de milliers d'autres molécules volatiles. Par ailleurs, le système olfactif voméronasal [A] possède environ 300 gènes codant des récepteurs olfactifs très spécifiques à certaines molécules olfactives, et n'a pas de projections corticales mais amygdaliennes et hypothalamiques. Cette organisation anatomo-fonctionnelle fait que le signal voméronasal est inconscient et provoque plutôt des effets endocriniens (par la régulation des hormones hypophysaires) ou comportementaux (agression, comportement maternel et sexuel). En général, la plupart des phéromones sont détectées par ce système, qui joue un rôle important dans le comportement sexuel (KEVERNE 2004). Le système olfactif principal [B] possède également des gènes codant pour des récepteurs spécifiques à des phéromones (LIBERLES & BUCK 2006), avec des circuits de projections spécifiques (BOEHM & al. 2005 ; projections non indiquées sur le schéma, car les données anatomiques actuellement disponibles sont contradictoires). Le système olfactif principal possède aussi et surtout environ un millier de gènes codant pour des récepteurs olfactifs permettant de détecter des milliers de molécules différentes et possède de nombreuses projections corticales, permettant ainsi un traitement conscient et plus cognitif de l'information olfactive. Bien qu'il puisse détecter des phéromones, qui sont des signaux souvent innés, ce système est en général plutôt impliqué dans le traitement des informations olfactives acquises. L'activation du système olfactif principal produirait principalement une analyse des molécules odorantes de l'environnement et l’association de ces stimuli olfactifs à des contextes.
Ainsi, les caractéristiques physicochimiques des molécules font qu'il existe des milliers de signaux élémentaires distincts les uns des autres et l'organisation structurelle des deux organes olfactifs permettent la détection de nombreux signaux olfactifs élémentaires et hautement spécifiques. Pour toutes ces raisons, le système olfactif est le système où la probabilité de trouver des signaux innés du comportement de reproduction est la plus grande.
Olfaction & comportement de reproduction
Par rapport au comportement de reproduction, une des principales fonctions des phéromones chez l'animal serait d'identifier le partenaire de sexe opposé (STOWERS & al. 2002 ; DULAC & TORELLO 2003). Mais, chez les primates de l'Ancien Monde (Catarhiniens), dont l'Homme, cette capacité aurait été perdue en raison de l'altération au cours de l'évolution du système voméronasal (ZHANG & WEBB 2003). Il est peu probable que cette discrimination hétérosexuelle soit réalisée chez les catarhiniens par d'autres phéromones, en raison d'une possible conservation phylogénétique des caractéristiques structurelles et fonctionnelles des phéromones (RAMPIN & al. 2006 ; KELLY 1996).
En cohérence avec cette détérioration du système voméronasal, et en cohérence avec l'important développement des structures télencéphaliques au détriment des structures olfactives, le résultat des différentes expériences réalisées chez l'Homme avec des phéromones potentielles est plutôt négatif (FOIDART & al. 1994 ; DESCOINS 2000 ; GOWER & RUPARELIA 1993). Par exemple, le système olfactif principal permet de reconnaître des liens de parenté, ce qui pourrait jouer un rôle dans le choix des partenaires sexuels. Mais cette capacité est acquise (PORTER & MOORE 1981). Un autre effet, mais physiologique et non comportemental, est la synchronisation des cycles menstruels des femmes qui vivent en communauté (WELLER & WELLER 1993). Mais certains chercheurs n'ont pas réussi à répliquer ce résultat (WILSON & al. 1991). D'autres résultats montrent une légère modification de l'état émotionnel, mais non spécifique à la reproduction. En conclusion de ces différentes études, on observe qu'aucune molécule testée n'a induit ou déclenché un comportement spécifique à la reproduction.
Les résultats les plus intéressants, par rapport au comportement de reproduction, concernent l’exposition aux androgènes, mais avec des résultats extrêmement contradictoires. On a ainsi montré que l'exposition expérimentale à l'androsténol augmente chez les femmes la durée de leur contact social avec les hommes (COWLEY & BROOKSBANK 1991). La molécule ne provoque pas le comportement de reproduction, mais elle permet de l'initier en favorisant le rapprochement hétérosexuel. Mais d'autres expériences ont montré que l'androsténone induit au contraire, chez les femmes, des réactions émotionnelles négatives envers les hommes (FILSINGER & al. 1985), que les hommes produisent simultanément les deux molécules, et que l'effet olfactif de l'androsténone est plus fort que celui de l'androsténol (MAIWORM 1990) : l'effet global, en situation "naturelle", devrait alors être répulsif.
Une autre expérimentation, présentée sous toute réserve, suggère qu'une phéromone naturellement sécrétée par l'homme augmente de manière significative les relations et les activités sexuelles des hommes : les rencontres non prévues avec des femmes ont augmenté de 35,3 %, le fait de dormir avec une femme a progressé de 35,3 %, les caresses et les baisers ont également progressé de 41,2 %, et la fréquence des relations sexuelles a augmenté de 47 %. Par contre, l'augmentation de 23,5 % de la fréquence de la masturbation n'est pas statistiquement significative (CUTLER & al. 1998). Ces résultats, qui ont été reproduit avec des femmes et des phéromones féminines (McCOY & PITINO 2002), suggèrent un effet facilitant ou attractant des phéromones sur les activités hétérosexuelles. Néanmoins, ces phéromones étant destinées à la commercialisation (GUEGUEN 2006 ; et cf. www.athenainstitute.com), en outre du problème de l'enjeux financier, il n'est pas précisé dans les articles quelles sont les molécules qui ont été utilisées. De plus, et surtout, comme ces molécules n'ont pas été testées ni avec des bisexuels et des homosexuels, ni avec des personnes ignorant tout de la sexualité, il est impossible de déterminer s'il s'agit d'un effet spécifique conduisant au coït vaginal hétérosexuel, ou plus simplement d'un effet émotionnel relativement général et de type euphorisant et/ou désinhibant (similaire par exemple à l'effet de désinhibition sociale et sexuelle de l'alcool).
Par ailleurs, on observe que chez les chimpanzés pan paniscus (Bonobo) près d'un quart des activités sexuelles sont réalisées par des individus prépubères (HASHIMOTO 1997 ; DE WAAL 1990). Ensuite – et surtout – dans certaines sociétés ou groupes humains sexuellement libéraux, les enfants ont des activités sexuelles dès l'âge de 3-4 ans. (SUGGS 1966 ; MALINOWSKI 1970 ; HENRY & HENRY 1974 ; KINSEY & al. 1948 ; MARTINSON 1994) (données détaillées à la section "1.a – Description des comportements de reproduction", page 3 ). Ces données suggèrent que, chez l'Homme, les éventuelles phéromones sexuelles (et également les hormones sexuelles), qui apparaissent bien plus tard, essentiellement à la puberté (BRUCE & COHN 1994), n'ont qu'un rôle secondaire dans l’apprentissage et l'expression de la sexualité et du comportement de reproduction.
En conclusion, en fonction des données éthologiques et expérimentales humaines déjà disponibles, on constate que les phéromones ont chez l'Homme un effet faible, essentiellement physiologique, avec quelques effets émotionnels qui modulent indirectement et faiblement des comportements en cours. Par rapport au comportement de reproduction, le seul effet comportemental des phéromones semblerait être, à partir de la puberté, de faciliter les interactions hétérosexuelles, favorisant ainsi indirectement la reproduction.
L'ensemble des données présentées semble suffisant pour conclure avec une bonne probabilité de validité qu'il n'existe pas chez l'Homme de processus olfactifs innés spécifiques du coït vaginal.
b.2 – Vision
La vision est un canal sensoriel majeur chez l'être humain. Certains auteurs ont donc supposé que des signaux visuels pourraient servir de signaux spécifiques au déclenchement du comportement de reproduction.
Les photons qui constituent le rayonnement électromagnétique et la lumière sont des particules d'énergie. Ils peuvent être distingués les uns des autres par leur longueur d'onde comprise entre 103 à 10-14 mètre. Les structures sensorielles de la vision sont les récepteurs (3 types de cônes et 1 type de bâtonnets) des cellules neurosensorielles de la rétine qui détectent les photons dont la longueur d'onde est comprise entre 400 à 700 nm. En fonction de ces caractéristiques, au mieux, à la lumière du jour, il n'est possible que de détecter trois signaux visuels élémentaires. De plus, au niveau anatomique, il ne semble pas exister chez les rongeurs de connexions entre les structures visuelles et les neurones de l'hypothalamus qui contrôlent le système hormonal de la reproduction (neurones à GnRH / LHRH – YOON & al. 2005). Pour ces raisons, il est peu probable qu'il existe un signal visuel élémentaire inné du comportement de reproduction.
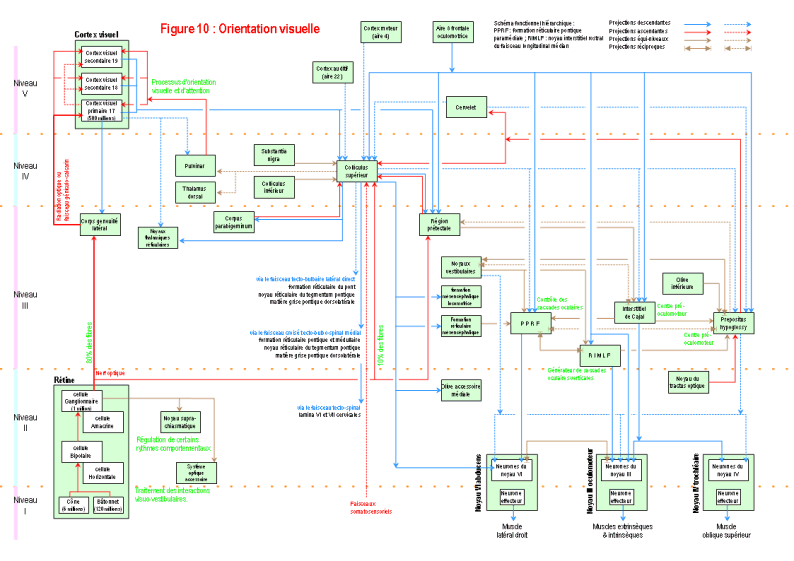
Néanmoins, il est possible qu'une information élaborée, extraite du traitement des stimuli visuels élémentaires, puisse servir de signal inné. Ce traitement pourrait avoir lieu dans les parties plus centrales du système visuel, en particulier dans les structures corticales.
Desmond Morris (1970) a formulé l'hypothèse que les fesses et les seins seraient des signaux visuels innés. D'après cette hypothèse, la similitude entre la forme des seins et des fesses s'explique par la nécessité, dans l’espèce humaine, et du fait du coït en face à face, de transposer un signal excitateur postérieur spécifique, les fesses, en un signal antérieur similaire, les seins. Les seins seraient ainsi des « fesses par devant » signal adaptatif excitateur spécifique à l’espèce humaine. Diverses enquêtes déclaratives et comportementales ont montré que les fesses et les seins étaient bien les deux signaux sexuels les plus attractifs pour les mâles de l’espèce humaine. Néanmoins, la question cruciale est de savoir si ces signaux sont innés ou bien s'ils sont acquis par des processus d'associations ou de conditionnements au cours des expériences sexuelles antérieures.
Les études génétiques, développementales, anatomiques et fonctionnelles du système visuel (voir Figure 10 pour l'orientation visuelle, et ROSENZWEIG & al. 2002 p. 281-321 pour le traitement cortical permettant la vision), montre que le système visuel est composé d'un grand nombre de petites structures (modules fonctionnels) qui ont chacune un rôle limité dans le traitement de l'information visuelle (traitement de la luminosité du signal par la rétine, génération des saccades oculaires par le noyau interstitiel rostral du faisceau longitudinal médian, traitement des couleurs par l'aire visuelle 4 ou des caractéristiques dynamiques (mouvement, direction, ...) par l'aire visuelle 5, etc.). C'est la coordination de toutes ces traitements élémentaires qui est à l'origine de la vision. En fonction de toutes ces connaissances actuelles, l'hypothèse de l'innéité d'une information visuelle ne semble guère plausible car il semble impossible d'expliquer le codage génétique (ou éventuellement épigénétique) et la localisation de l'image prototypique préformée à laquelle cette information visuelle complexe doit être comparée. En effet, la reconnaissance des formes complexes est située dans les aires corticales associatives, qui sont les régions les plus complexes du cerveau ( Figure 1, niveau [7], page 14 ). Or, par rapport au développement, seules les grandes voies de connexion du système visuel semblent être codées génétiquement, et de nombreuses capacités du système visuel se développent en interaction avec l'environnement. De plus, un gène code une protéine, qui, dans le meilleur des cas, ne peut servir que de molécule de guidage global aux connexions synaptiques. Comment alors expliquer le codage spécifique et précis des milliards de synapses nerveuses vraisemblablement nécessaires à l'interprétation des formes complexes des fesses et des seins dans les aires associatives du cortex temporal inférieur (GAZZANIGA & al. 2002, p. 195-204) ? Comment expliquer la stabilité de ce codage dans des régions connues pour être le siège de phénomènes de plasticité, de stabilisation sélective et de réorganisation ? Comment enfin ce signal participerait-il à l'exécution du coït vaginal fécondant ?
Par ailleurs, chez les mammifères sub-primates, ce sont principalement des signaux chimiques qui contrôlent la reproduction, au sein de structures neurales phylogénétiquement anciennes. Ce canal de communication semble le mieux adapté aux réalités biologiques des organismes mammaliens : il suffit de quelques molécules, de quelques récepteurs et d'un simple réseau neural pour distinguer les partenaires, activer des voies sensorielles, motrices, et déclencher des séquences comportementales. Pour quelles raisons évolutives ce système simple et efficace aurait-il été remplacé par un système complexe de traitement de l'information visuelle au sein des régions néocorticales les plus phylogénétiquement récentes ?
De plus – et surtout – sachant que l'Homme n'a que quelques pour cent de différence génétique avec les autres primates, il est quasiment impossible, en fonction des connaissances actuelles, de concevoir comment des mutations ou des modifications génétiques aléatoires dans un organisme protohumain et sur un nombre restreint de gènes peuvent avoir codé par anticipation une image prototypique de la nouvelle morphologie finale d'homo sapiens. Ce phénomène est, en fonction des connaissances actuelles, rigoureusement impossible.
D'autres auteurs ont proposé que chez les hominoïdes et les singes de l'ancien monde le système voméronasal aurait été remplacé par un système visuel simple, basé sur la perception chromatique, et qui permettrait par exemple de détecter les modifications de couleur de la peau sexuelle des femelles de certains primates (ZHANG & WEBB 2003). Néanmoins, même si une couleur est un signal simple, il semble nécessaire d'associer à cette détection chromatique un traitement cognitif plus élaboré (par exemple une analyse de la forme), afin d'éviter que les réactions instinctuelles ne se déclenchent à chaque perception dans l'environnement du signal chromatique inné. De surcroît, en plus du caractère hypothétique de ce circuit visuel, une expérimentation chez le macaque rhésus, qui effectivement ne semble plus avoir de système voméronasal fonctionnel (GOLDFOOT 1981), montre que la vision, au contraire de l'olfaction, n'est pas un mode sensoriel de communication des signaux sexuels innés. Des mâles sexuellement naïfs et privés expérimentalement de leur odorat n'engagent aucune activité sexuelle bien qu'ils voient une femelle réceptive ayant de surcroît le sexe tumescent et coloré (MICHAEL & KEVERNE 1968).
En conclusion, dans l'état actuel des connaissances, les photons ne peuvent véhiculer une information sexuelle élémentaire, et il n’y a pas, chez l'Homme, de signal visuel inné qui provoquerait (ou au minimum qui induirait) le coït vaginal reproducteur.
b.3 – Audition
Introduction : processus auditifs
Les structures sensorielles de
l'audition sont des cellules épithéliales ciliés sensorielles de l'organe
spiral de la cochlée qui détectent les mouvements provoqués par les vibrations
moléculaires de l'air ou de l'eau. Les vibrations moléculaires n'ayant pas de
caractéristiques sexuelles spécifiques, il ne peut donc exister de signal
auditif élémentaire inné du comportement de reproduction.
De plus, au niveau anatomique, il ne semble pas exister chez les rongeurs de connexions entre les structures auditives et les neurones de l'hypothalamus qui contrôlent le système hormonal de la reproduction (neurones à GnRH / LHRH – YOON & al. 2005).
Néanmoins, il est possible qu'une information élaborée, extraite du traitement des stimuli auditifs élémentaires, puisse servir de signal inné. Ce traitement pourrait avoir lieu dans les parties plus centrales du système auditif. L'organisation anatomique du système auditif est donné à la Figure 11 (pour des explications détaillées, voir : NIEUWENHUYS & al. 1988 ; BOSSY 1990 ; GUYTON 1989). De manière similaire au système visuel, le système auditif est composé d'un grand nombre de structures (noyau cochléaire dorsal, noyau olivaire supérieur, colliculus inférieur ...) qui ont chacune un rôle limité dans le traitement de l'information auditive (accommodation auditive, initiation des réactions auditives réflexes (réflexe stapédien par exemple), localisation de la source sonore, etc.).
En fonction de ces données anatomiques et fonctionnelles, une information auditive préformée spécifique du comportement de reproduction devrait être recherchée plutôt au niveau des cortex auditifs. Mais des problèmes similaires à ceux déjà exposés pour le système visuel (milliards de synapses, développement épigénétique, réorganisation corticale, etc., voir section précédente) rendent peu probable l'existence d'un signal auditif préformé, spécifique du comportement de reproduction.
Audition & comportement de reproduction
Par rapport aux données connues, on observe que les rats émettent des ultrasons, qui ont la caractéristique d'un chant (HOLY & GUO 2005), durant la phase appétitive du comportement sexuel (BARFIELD & al. 1979). Mais ces vocalisations sont apprises (MAGGIO & al. 1983) et sont également émises durant le comportement d'agression, et surtout durant les jeux et les chatouilles. L'analyse des différents contextes où ces vocalisations sont émises suggère qu'elles seraient provoquées par des états affectifs positifs (BURGDORF & PANKSEPP 2001), et qu'elles ne seraient donc pas des signaux auditifs spécifiques du comportement de reproduction.
L'Homme émet également des vocalisations durant les activités érotiques, mais elles ne semblent pas être des signaux spécifiques de la reproduction. Elles sont plutôt une conséquence physiologique provoquée par l'état de plaisir intense. On observe d'ailleurs qu'il s'agit d'une caractéristique générale et que tous les états affectifs intenses provoquent des réactions motrices et vocales particulières.
En conclusion, dans l'état actuel des connaissances, les vibrations moléculaires ne peuvent véhiculer une information sexuelle élémentaire, et on ne connaît pas de données expérimentales ou éthologiques montrant qu'il existe chez les mammifères, et surtout chez l'Homme, des signaux auditifs innés, spécifiques du comportement de reproduction.
b.4 – Toucher
Existe-t-il des signaux somatosensoriels innés et spécifiques, participant au contrôle du comportement de reproduction ?
Introduction : processus somatosensoriels
Le système somatosensoriel est constitué de plusieurs types de récepteurs (mécanorécepteurs, nocicepteurs, thermorécepteurs ...) localisés dans la peaux, les viscères, les muscles et les tendons de l'organisme (Figure 12). À la différence des systèmes sensoriels où les récepteurs sont localisés dans des organes sensoriels, les différents récepteurs somatosensoriels sont disséminés dans tout le corps. Ces récepteurs permettent la détection de stimuli nociceptifs, de la température, de l'étirement, de la pression, des vibrations ou du contact. À ce niveau, on remarque qu'aucun de ces récepteurs ne peut être spécifique d'un signal sexuel. Tout contact avec le corps active des récepteurs somatosensoriels, que ce contact soit sexuel ou non. Il n'existe donc pas de signaux somatosensoriels élémentaires spécifiques du comportement de reproduction.
Ces différents récepteurs somatosensoriels projettent des fibres sensitives dans la moelle épinière (Figure 13) où peuvent être organisé des réflexes moteurs. À ce niveau peut exister un précablage inné qui pourrait provoquer des réactions motrices réflexes spécifiques du comportement de la reproduction.
De la moelle épinière partent également des faisceaux sensoriels qui transmettent au différents niveaux du système nerveux les informations somatosensorielles (Figure 14). La plupart de ces faisceaux se projettent dans des structures précises du cerveau (voir l'exemple du faisceau spino-thalamique à la Figure 15). On remarque que le système somatosensoriel est en fait constitué de systèmes plus ou moins distincts. Ces systèmes sont constitués par des combinaisons de certains types de récepteurs, localisés en des régions particulières du corps (pulpe des doigts, peau glabre ou poilue, tissu muco-cutané, etc.), et connecté par des liaisons spécifiques avec certaines structures du système nerveux (motoneurone médullaire, substance réticulée, thalamus intralaminaire ou cortex somatosensoriel, etc.). Cette organisation permet l'émergence de propriétés particulières (réflexes, douleur, tact protopathique ou épicritique, discrimination fine ou grossière, etc.) (pour des explications détaillées, voir : NIEUWENHUYS & al. 1988 ; WILLIS & COGGESHALL 1991 ; BESSON & al. 1994 ; BOSSY 1990 ; MARIEB 1993). C'est à ce niveau qu'il pourrait exister un système particulier du système somatosensoriel, inné, comme éventuellement celui des sensations corporelles agréables (OLAUSSON & al. 2002 ; WESSBERG & al. 2003) ou érotiques, et qui pourrait être impliqué dans le comportement de reproduction.
Système somatosensoriel & comportement de reproduction
Par rapport au comportement sexuel, on remarque au niveau anatomique qu'il existe chez les rongeurs des connexions entre les structures somatosensorielles et les neurones de l'hypothalamus qui contrôlent le système hormonal de la reproduction (neurones à GnRH / LHRH – YOON & al. 2005).
On observe par ailleurs, en particulier chez les rongeurs et les félidés, une activité spécifique de la reproduction qui est contrôlée par des signaux tactiles. Chez la femelle, en période œstrale, la lordose (décrite plus en détail à la section traitant des réflexes innés, "1.d.g.1 – Réflexes spécifique au comportement de reproduction", p. 26) est déclenchée par des stimuli tactiles au niveau des flancs. Mais il convient de noter que ce signal tactile n'est pas spécifique de la reproduction : n'importe quel contact avec les flancs déclenche ce réflexe postural. En conclusion, c'est la posture de lordose qui est innée et spécifique de la reproduction, mais pas le signal somatosensoriel qui la déclenche. Par ailleurs, la lordose n'est pas observée chez la femme, où d'ailleurs elle n'aurait que peu d'intérêt, n'étant guère adaptée à la position d'accouplement des humains généralement en face à face en raison de la position plus antérieure du vagin dans l'espèce humaine.

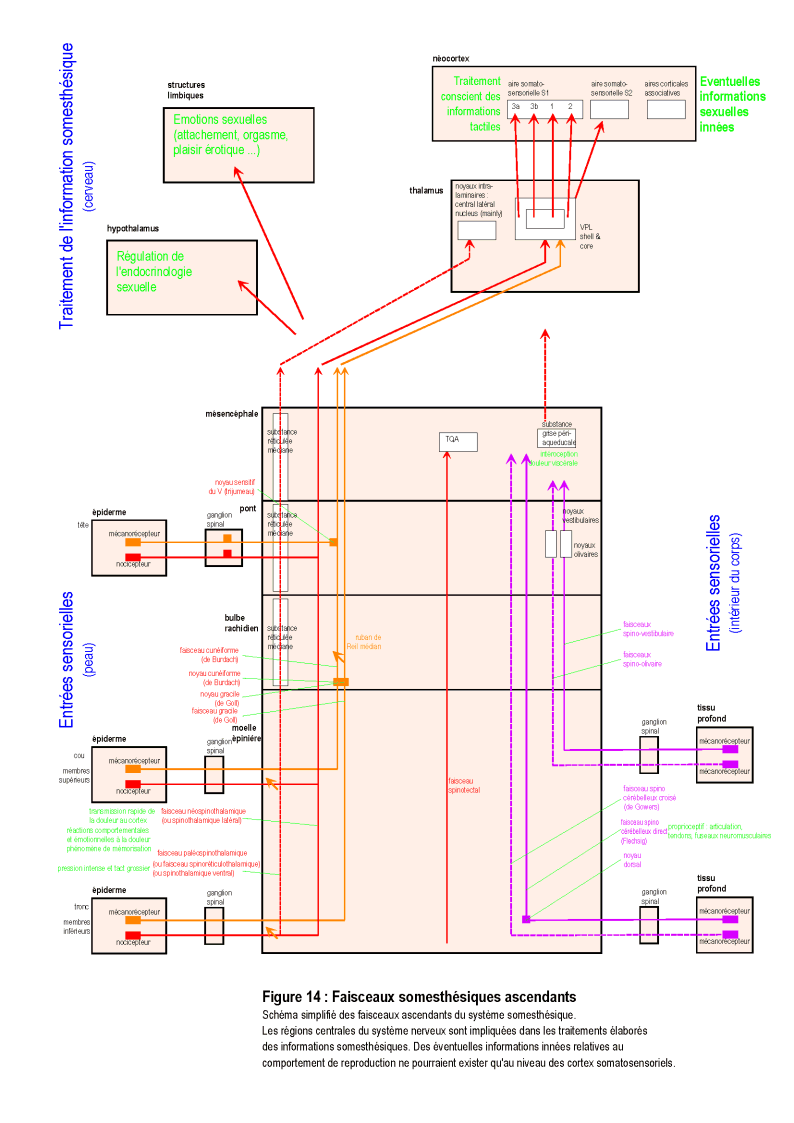
Plus intéressant, on observe que la stimulation mécanique du pénis ou du clitoris provoque l'éjaculation [1], phénomène physiologique spécifique à la reproduction et qui ne peut apparemment être déclenché, au moins initialement, que par des stimuli somatosensoriels. Néanmoins, là encore, il convient de noter que ce signal somatosensoriel n'est pas spécifique de la reproduction : de nombreux types de contacts avec le pénis, distincts de ceux particuliers au coït vaginal, peuvent provoquer l'éjaculation. En conclusion, c'est l'éjaculation qui est innée et spécifique de la reproduction, mais pas le signal somatosensoriel qui la déclenche. Pour donner un exemple théorique, un signal somatosensoriel inné et spécifique de la reproduction aurait pu être le déclenchement de l'éjaculation après une analyse corticale des sensations péniennes produites par l'intromission, lorsque cette analyse aurait abouti à la reconnaissance de la forme du vagin par comparaison au niveau des aires somatosensorielles avec une éventuelle image préformée de cet organe génital. Mais aucune donnée connue ne corrobore cette possibilité.
En conclusion, dans l'état actuel des connaissances, les stimulations somatosensorielles n'ont pas de spécificités sexuelles élémentaires, et on ne connaît pas de données expérimentales ou éthologiques montrant qu'il existe chez les mammifères et chez l'Homme des signaux somatosensoriels innés et spécifiques du comportement de reproduction.
b.5 – Goût
Existe-t-il des signaux gustatifs innés et spécifiques, participant au contrôle du comportement de reproduction ?
Introduction : processus gustatifs
Les structures sensorielles de la gustation sont les cellules épithéliales ciliés sensorielles des bourgeons gustatifs qui détectent les substances chimiques présentes dans la cavité buccale. Les molécules gustatives peuvent avoir de nombreuses caractéristiques physicochimiques qui les rendent distinctes et particulières des autres. Elles peuvent provenir et être spécifiques d'un autre organisme et pourraient être un signal spécifiquement sexuel. Néanmoins la structure et l'organisation des récepteurs gustatifs font que seul cinq modalités sensorielles sont détectées : salé, sucré, acide, amer et umami. Ces modalités n'ayant pas de caractéristiques sexuelles spécifiques, il ne peut donc exister de signal gustatif élémentaire inné du comportement de reproduction.
Il est néanmoins possible qu'une information extraite du traitement des stimuli gustatifs élémentaires puissent servir de signal inné. Ce traitement pourrait avoir lieu dans les parties plus centrales du système gustatif, qui s'étend jusqu'au cortex. L'organisation anatomique du système gustatif est donné à la Figure 16 (pour des explications détaillées, voir : NIEUWENHUYS & al. 1988 ; BOSSY 1990 ; GUYTON 1989). De manière similaire aux autres systèmes sensoriels, le système gustatif est composé de plusieurs petites structures qui ont chacune un rôle limité dans le traitement de l'information gustative (déclenchement de la sécrétion de salive ou de suc gastrique, alliesthésie, etc.). Néanmoins, plus encore que pour le système visuel ou auditif, l'existence d'information gustative préformée spécifique au comportement de reproduction semble peut probable.
Gustation & comportement de reproduction
Par rapport aux données connues, on observe chez certains mammifères des activités de léchage dans les activités qui précédent la copulation. Mais ces activités sont phéromones-dépendantes et n'ont pas comme finalité le recueil de molécules gustatives spécifiques de la reproduction. Les phéromones ainsi recueillies passent dans l'organe voméronasal par une voie de communication entre les cavités buccale et nasale (canal nasopalatin, chez les carnivores, les ongulés, les insectivores et les primates du Nouveau-Monde). Il ne s'agit donc pas de signaux gustatifs.
En conclusion, dans l'état actuel des connaissances, les récepteurs gustatifs ne peuvent transmettre une information sexuelle élémentaire, et on ne connaît pas de données expérimentales ou éthologiques montrant qu'il existe chez les mammifères et chez l'Homme des signaux gustatifs innés de la reproduction.
b.6 – Conclusion relative aux systèmes sensoriels
En fonction des données présentées dans les sections précédentes, le seul système sensoriel où il existe des signaux élémentaires qui peuvent avoir une spécificité sexuelle et où les récepteurs peuvent détecter de manière innée cette spécificité est le système olfactif. Pour ces raisons, les seuls signaux élémentaires innés et spécifiques de la reproduction actuellement connus sont les phéromones sexuelles. Quant aux informations "préformées" qui pourraient théoriquement exister dans les structures plus centrales des systèmes sensoriels, et ainsi permettre la reconnaissance innée d'une information sensorielle spécifique de la reproduction, aucune donnée expérimentale ne corrobore leur existence.
En conclusion, les phéromones sont un mode de communication majeur pour de nombreuses espèces animales, mais elles ne jouent qu'un rôle mineur dans le comportement de reproduction humain. Elles ne sont pas le facteur à l'origine des différentes activités motrices qui permettent le coït vaginal reproducteur.
g – Système moteur
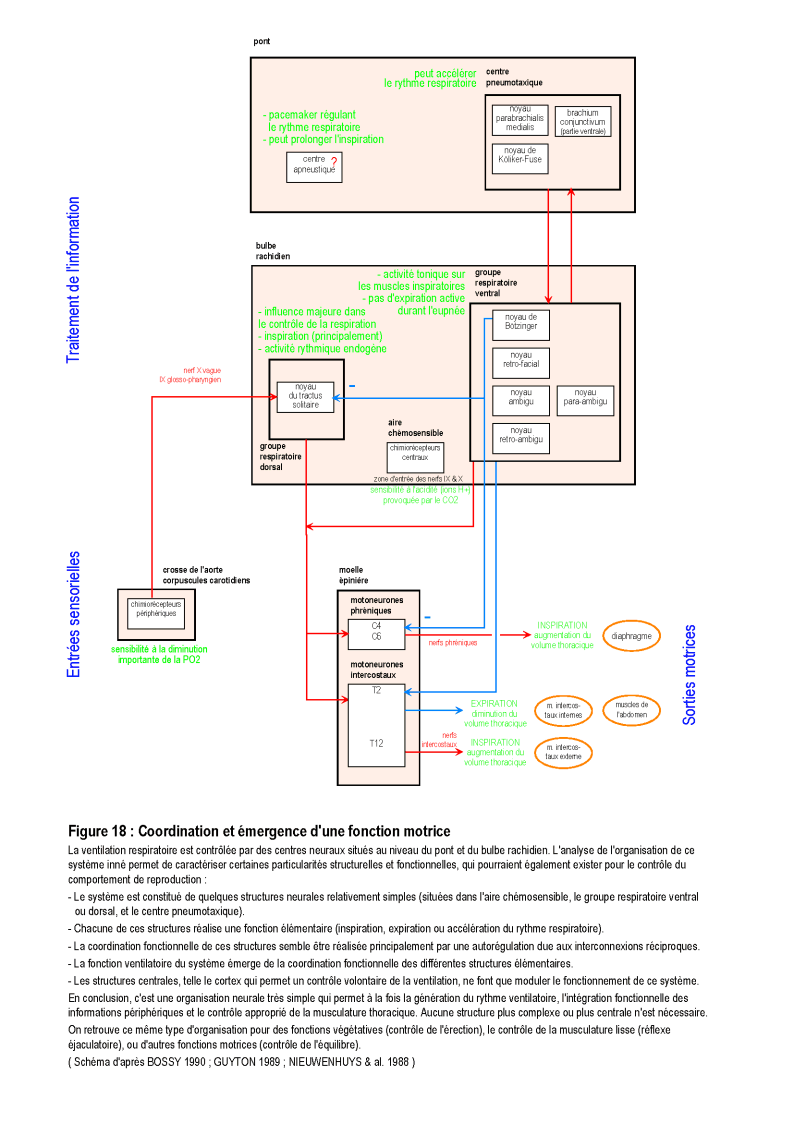
On devrait pouvoir trouver des structures neurales organisées de telle manière qu'elles peuvent initier, exécuter et contrôler sans aucun apprentissage des séquences motrices spécifiques permettant le coït vaginal.
g.1 – Réflexes spécifiques au comportement de reproduction
Au niveau d'organisation du système nerveux immédiatement supérieur aux cellules sensorielles, apparaissent les mécanismes réflexes (LAGET 2005 ; Figure 1, niveau [3], page 14 ) et les mouvements coordonnés plurisegementaires et suprasegmentaires (BUSER 2005 ; Figure 1, niveau [4] ).
Introduction : processus moteurs
Dans un premier temps, quelles sont les connaissances actuelles relatives à des systèmes moteurs innés et bien étudiés, et que peuvent elles apporter dans la recherche de l'existence d'un instinct du comportement de reproduction ?
Par rapport à l'activité réflexe, les études des réflexes spinaux (réflexe myotatique, de flexion, d'extension contralatéral ...) montrent qu'il suffit d'un neurone sensoriel connecté à un neurone moteur pour faire apparaître un réflexe moteur. Par ailleurs, les études du contrôle des mouvements coordonnés du système digestif des crustacés montrent que l'interconnexion en réseau de quelques dizaines de neurones ayant des propriétés bioélectriques différentes est suffisant pour faire émerger des séquences motrices adaptées et relativement élaborées (Figure 17). En synthèse, on observe dans ces deux organisations neurales simples que l'interconnexion hautement spécifique entre des neurones sensoriels, moteurs et quelques dizaines d'interneurones organisés en réseaux permet l'émergence de réflexes moteurs adaptés ou de mouvements coordonnés élémentaires. Ces données indiquent qu'il serait possible que certaines séquences motrices du comportement de reproduction soient contrôlées de manière innée juste par quelques neurones qui peuvent se trouver même au niveau médullaire. Ce pourrait être le cas par exemple des séquences de l'intromission du pénis et des poussées pelviennes rythmiques.
Par rapport à un contrôle un peu plus élaboré d'activités motrices, l'étude par exemple de la ventilation pulmonaire permet d'apporter des informations complémentaires (Figure 18) (pour des explications détaillées concernant la ventilation, voir : MARIEB 1993 ; BOSSY 1990 ; GUYTON 1989 ; NIEUWENHUYS & al. 1988 ). Cet exemple met en exergue les caractéristiques générales des réflexes élaborés, à savoir une organisation en réseau de structures neurales plutôt simples, qui chacune réalise une fonction élémentaire, et situées à proximité des récepteurs et des effecteurs. Bien qu'il existe des connexions avec des structures centrales plus complexes, ces connexions n'ont apparemment qu'un rôle modulateur ou coordonnateur. L'analyse fonctionnelle de ces processus neuraux, et d'autres similaires tel l'équilibre, suggère que le contrôle moteur est décentralisé : le niveau local effectue un contrôle précis du réflexe, le niveau plus central réalise une régulation et une coordination plus globale, en fonction d'informations internes et externes à l'organisme.
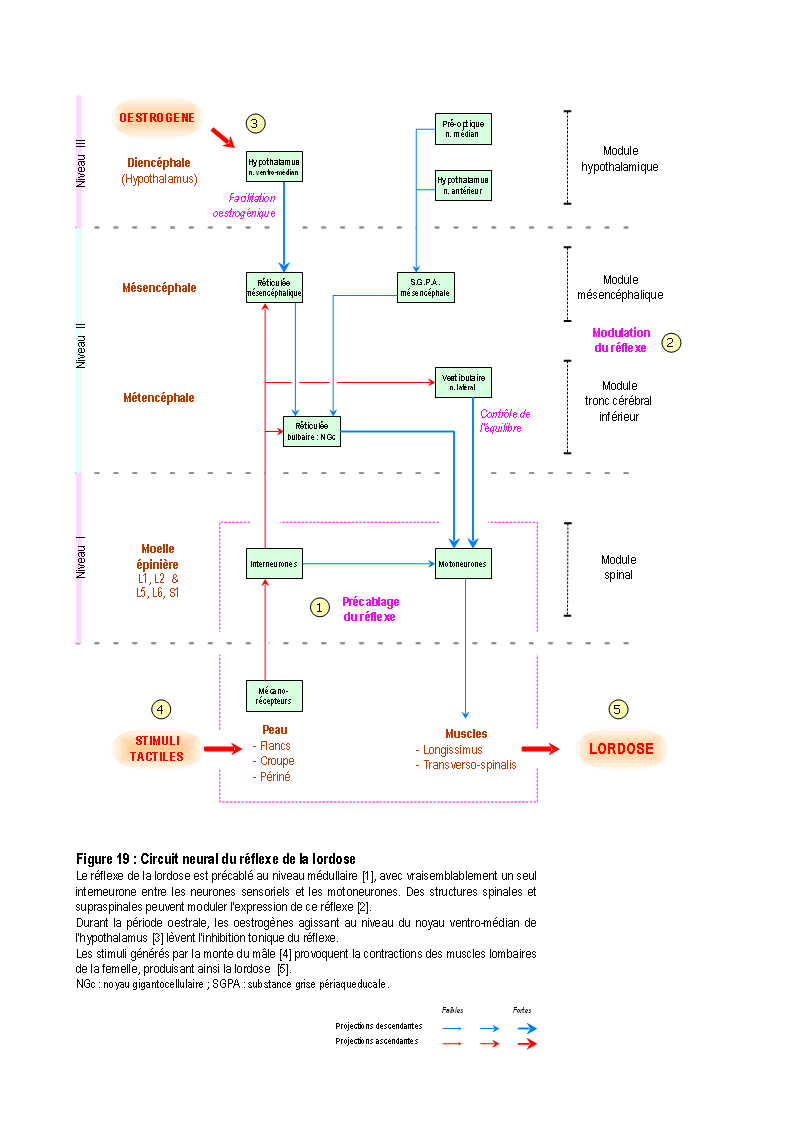
En conclusion, il apparaît que des séquences motrices réflexes de la reproduction pourraient être codées au niveau médullaire par quelque dizaine de neurone, organisé en réseau. Il ne semble pas nécessaire que le comportement de reproduction soit entièrement "programmé" dans une structure centrale relativement complexe, telle par exemple l'aire préoptique médiane de l'hypothalamus. S'il existe des séquences réflexes innées relatives au comportement de reproduction, elles doivent certainement être recherchés dans la coordination de petites structures neurales situées à un ou deux relais neuronaux des neurones sensoriels ou moteurs impliqués dans les séquences motrices de la reproduction.
Système moteur & comportement de reproduction
Chez les mammifères inférieurs, on observe le développement et l'existence, généralement sous l'effet des hormones sexuelles, de structures précablées permettant l'exécution de réflexes spécifiques, tel par exemple la lordose ou les poussées pelviennes permettant l'intromission. La lordose lombaire est sans doute le meilleur exemple actuellement connu de séquence motrice instinctuelle spécifique de la reproduction (Figure 19). La lordose est un réflexe somato-sensoriel œstrogène-dépendant, précablé au niveau médullaire et mésencéphalique, et sous contrôle de l'hypothalamus. Le précablage est relativement simple, au niveau des segments médullaires T11 à L6, avec vraisemblablement un seul interneurone qui reçoit des influx mésencéphaliques modulateurs, œstrogène-dépendant. Les récepteurs sensoriels sensibles à la pression, situés dans le derme des flancs de l'animal, transmettent des signaux tactiles, qui, en l'absence d'inhibition des interneurones, sont directement transmis aux motoneurones des muscles du pelvis et de la colonne vertébrale. Une pression sur les flancs de la femelle, en général au cours de la monte, provoque la courbure du dos et l'élévation de la croupe, présentant ainsi le périnée, ce qui facilite la copulation (PFAFF & al. 1994). À noter que si ce réflexe inné est spécifique de la reproduction, les stimulations tactiles qui le déclenchent ne le sont pas. De nombreux types de contacts tactiles avec les flancs provoquent la lordose.
Chez l'Homme, par rapport aux structures précablées, on ne connaît que des structures neurales innées qui contrôlent des phénomènes physiologiques directement impliqués dans la reproduction, comme par exemple le circuit réflexe de l'éjaculation ou de l'érection (nerf dorsal du pénis, noyaux sensoriels lombo-sacrés, noyaux parasympathiques sacrés et noyaux d'Onuf, nerfs pelvien et honteux – GIULIANO & RAMPIN 2004), mais aucune structure qui contrôlerait des aspects moteurs, comme le positionnement correct préalable à l'intromission. À noter également que si ces réflexes physiologiques innés sont spécifiques de la reproduction, les stimulations qui les déclenchent ne le sont pas : de nombreux types de stimulations tactiles du pénis provoquent l'érection et l'éjaculation (cf. également la section b.4 – "Toucher", page 22). La seule séquence motrice instinctuelle spécifique de la reproduction, sous réserve de vérification expérimentale, pourrait être les poussées pelviennes.
Les observations comportementales confirment cette absence de précablage moteur spécifique à la reproduction. À partir de quelques cas où, à l'âge adulte, des humains, hommes ou femmes, n'ont pu accéder à la connaissance du coït vaginal, on a remarqué qu’en général, au cours de leurs activités sexuelles, ils ne pratiquaient pas le coït vaginal, même lorsqu'ils sont l'un contre l'autre (RICHARD-FOY 2004 ; voir également l'étude présentée à l'annexe IV, page 205) : il ne semble donc pas exister de mécanismes réflexes cruciaux provoquant les séquences motrices permettant l'intromission du pénis dans le vagin.
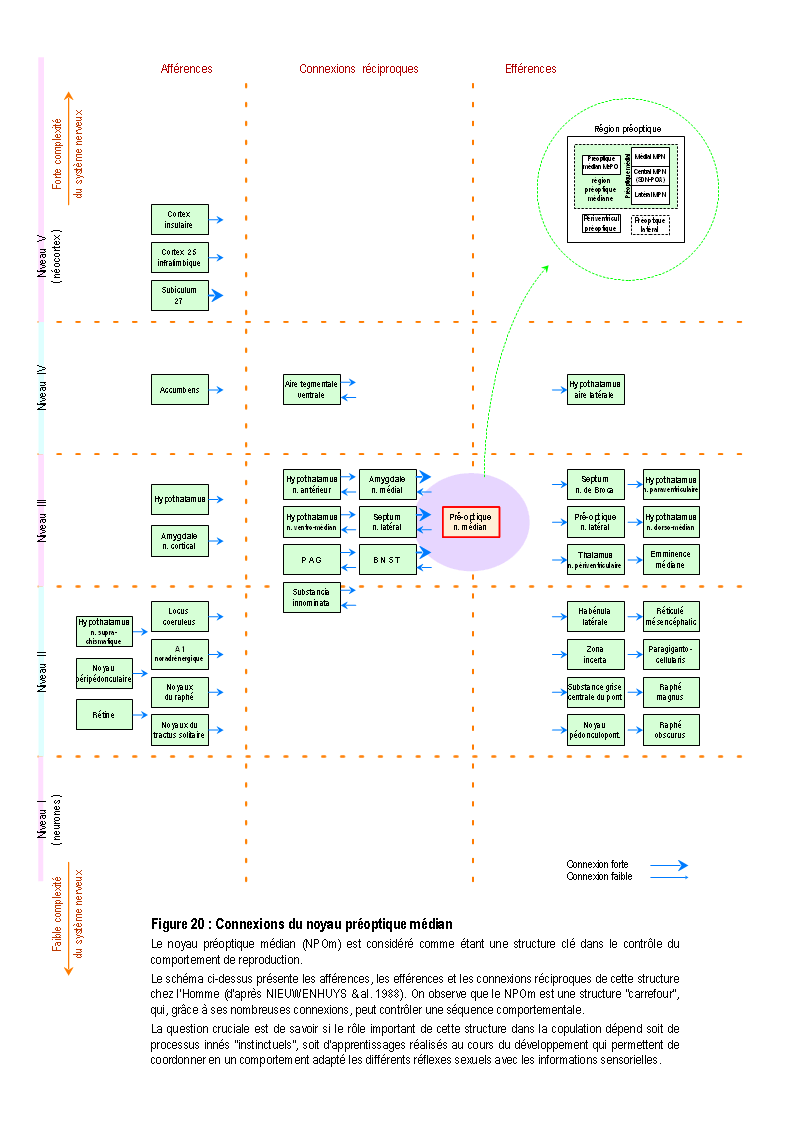
De plus, ces caractéristiques motrices innées élémentaires n'interviennent que dans le contrôle local du coït, et ne constituent que la partie finale du comportement de reproduction. Même si les poussées pelviennes étaient innées, cette séquence motrice ne peut à elle seule permettre la reproduction.
En conclusion, il existe chez les rongeurs des éléments moteurs instinctuels, mais ils ne concernent que des réflexes élémentaires, certes important pour la réalisation du comportement de reproduction, mais ne permettant pas sans un apprentissage qui les coordonne dans une organisation fonctionnelle supérieure, la réalisation d'un comportement cohérent et finalisé. Chez l'Homme, hormis peut-être le réflexe des poussées pelviennes, il ne semble exister aucun précablage moteur spécifique de la reproduction.
Figure 20
g.2 – "Programmation" comportementale
À un niveau d'organisation encore supérieur du système nerveux, apparaissent des structures capables de contrôler et de réguler de manière globale tout un ensemble d'informations et de séquences motrices ou autonomes ( Figure 1, niveau [5], page 14 ).
Il est possible d'envisager que le caractère inné du comportement de reproduction soit intégré à ce niveau supérieur, dans des structures qui soit permettent la coordination globale des différents réflexes sexuels élémentaires en un comportement de reproduction, soit contiennent « un programme central organisateur de la succession des opérations impliquées par les divers stades du mouvement (dès lors, l'acte n'est plus une succession de réflexes mais bien une séquence prédéterminée des étapes de ce mouvement) » (BUSER 2005).
Un des meilleurs candidats actuels de ce type de structure serait le noyau hypothalamique préoptique médian (NPOm) (Figure 20 ; NIEUWENHUYS & al. 1988). La lésion de cette structure, qui présente un dimorphisme sexuel et qui possède des relations avec la région génitale et le système voméronasal, supprime la copulation chez les rongeurs (MEISEL & SACHS 1994). Il est supposé que la fonction de cette structure serait soit 1) la régulation des aspects moteurs de la copulation ; soit 2) la régulation de la motivation sexuelle ; soit 3) la régulation de ces deux aspects, moteur et motivationnel (PAREDES 2003).
Pour certains auteurs, la lésions du NPOm chez les rongeurs mâles supprimerait l'aspect moteur de la copulation (EVERITT 1990) et pour d'autres l'aspect motivationnel (PAREDES 2003). Anatomiquement, le NPOm reçoit des afférences du système voméronasal, via l'amygdale et le noyau du lit de la strie terminale (MEREDITH 1991), des afférences provenant des organes génitaux au travers de relais spinaux et bulbaires (MALLICK & al. 1994), et se trouve à proximité du striatum ventral, qui est impliqué dans certains aspects de la motivation sexuelle (EVERITT 1990). La mesure de l'activité neuronale du NPOm lors de la copulation (SHIMURA & al. 1994), suggère, conformément aux connections anatomiques, que cette structure reçoit des informations phéromonales et participe à des aspects moteurs de la copulation, vraisemblablement par des facilitations des réflexes sexuels (DOMINGUEZ & HULL 2005). D'où, le NPOm ayant une position anatomique et fonctionnelle "carrefour", il est cohérent que sa lésion entraîne les effets observés.
Ces données provenant essentiellement des rongeurs, à défaut de pouvoir préciser les fonctions exactes du NPOm, permettent néanmoins de répondre à la question de l'innéité. On observe que c'est la séquence motrice de la monte qui est la plus perturbée par la lésion du NPOm (EVERITT 1990), séquence dont nous avons vu précédemment qu'elle serait apprise (les animaux mâles isolés depuis la naissance ne parviennent pas à copuler, cf. section "1.d – Le comportement de reproduction est-il inné ?", page 13). D'où, si la fonction de NPOm est le contrôle moteur de la copulation, il s'agit donc d'une capacité apprise et non innée. Dans ce cas, NPOm ne contiendrait pas un programme central organisateur inné, mais des modifications neurales acquises durant la période du développement, permettant le contrôle de la séquence de la monte à partir vraisemblablement de l'intégration d'informations provenant des nombreuses structures auxquelles NPOm est connecté. Si la fonction du NPOm est la motivation sexuelle, innée, qui est apparemment induite par les stimuli phéromonaux ou hormonaux, il s'agit alors d'une motivation innée phéromone-dépendante ou hormone-dépendante. Or, non seulement le système voméronasal est vestigial chez les hominoïdes et les primates de l'ancien monde (ZHANG & WEBB 2003), mais ses effets fonctionnels sont faibles et ne permettent pas la réalisation du coït vaginal (cf. section "1.d.b.1 – Olfaction", page 17) ; quant aux hormones, leur influence décroît avec le développement cortical (BUVAT 1996). Dans ce cas, la motivation sexuelle serait innée, mais uniquement chez les mammifères inférieurs puisque les mécanismes innés disparaîtraient parallèlement à l'évolution corticale.
En conclusion, actuellement, on ne connaît aucun exemple chez l'Homme d'une organisation neurale spécifique qui, de part son architecture spécifique, soit permet la coordination innée des réflexes sexuels élémentaires en un comportement de reproduction, soit contient le "programme" d'un ensemble de séquences motrices, planifiées, coordonnés et finalisés, permettant la réalisation innée et adaptée de l'ensemble (ou de la majeure partie) du comportement de reproduction.
Les seuls exemples connus de séquences motrices "programmées" ou "précablées" sont les réflexes sexuels élémentaires, telle la lordose, qui ont été décrits à la section précédente.
d – Régulation hormonale
En théorie, les hormones sont des facteurs parfaitement appropriés pour effectuer des régulations globales. La libération d'hormones, qui peuvent être diffusées dans tout l'organisme, peut permettre d'initier, d'inhiber, de moduler ou de coordonner de manière synchrone différents types de processus dans différentes régions du corps. En fonction des données provenant de l'expérimentation animale, ce type de régulation existerait au niveau d'un système constitué par l'hypothalamus, l'hypophyse et l'épiphyse. Chez les mammifères inférieurs ces structures neurales régulent la puberté et deux cycles physiologiques, saisonnier et œstral, qui interviennent dans la reproduction et le comportement sexuel.
Cycle saisonnier
Le contrôle saisonnier de la reproduction, observé dans de nombreuses espèces de mammifères (caprins, ovins et la plupart des espèces sauvages, sauf les primates), permet d'améliorer la survie de l'espèce par des naissances ayant lieu durant la période la plus favorable de l'année. Ce cycle saisonnier dépend du système rétine / hypothalamus / épiphyse / mélatonine, qui détecte les variations annuelles de la durée des jours. L'information photopériodique, perçue au niveau de la rétine, est transmise par l'intermédiaire de plusieurs relais nerveux (noyaux suprachiasmatiques (LEAK & MOORE 1997), ganglions cervicaux supérieurs ...) à l'épiphyse. À ce niveau, le cycle lumineux est traduit en rythme circadien de sécrétion de mélatonine. Cette hormone régule les hormones sexuelles par son action, suivant les espèces, soit au niveau hypothalamique sur la sécrétion de GnRH ou de LH, soit parfois par une action directe sur les gonades. Mais chez l'être humain, l'activité sexuelle et la reproduction existent tout au long de l'année et on n'observe, seulement dans les pays nordiques, qu'un effet mineur des saisons sur le comportement de reproduction (ALEANDRI & al. 1996 ; PANDI-PERUMAL & al. 2006).
Cycle œstral
Le contrôle œstral de la reproduction, qui dépend de l'axe hypothalamo-hypophysaire et des hormones sexuelles, permet de synchroniser la copulation avec l'ovulation. Mais contrairement à beaucoup de mammifères femelles, la femme a des activités sexuelles sur l'ensemble du cycle œstral. Néanmoins, on observe une augmentation de la fréquence du coït vaginal durant la période de fertilité, juste avant l'ovulation (WILCOX & al. 2004). Cependant, cet effet hormonal est relativement faible, et, surtout, il est général : toutes les activités sexuelles sont augmentées, masturbatoires ou avec un partenaire, hétérosexuelles ou homosexuelles (BURLESON & al. 2002). Cet effet hormonal, de type motivationnel, n'est donc ni spécifique de l'hétérosexualité, ni du coït vaginal.
Puberté
Quant au phénomène de la puberté, qui dépend d'une horloge biologique, des neurones à GnRH et des hormones sexuelles (SISK & FOSTER 2004), il permet d'initier de manière synchrone le comportement de reproduction avec la maturation physiologique de l'appareil reproducteur. Néanmoins, chez l'Homme, les effets de la puberté ne semblent pas être déterminant pour le comportement de reproduction.
En effet, on observe vers l'âge de 10 ans lors de la sécrétion des hormones surrénales (adrénarche), que la déhydroépiandrosterone (DHEA) a un effet sur l'attraction sexuelle. Néanmoins cet effet est faible et l'attraction sexuelle n'est pas spécifique au sexe opposé (HERDT & MCCLINTOCK 2000 ; MCCLINTOCK & HERDT 1996). Cet effet mineur ne peut donc pas être considéré comme étant un facteur participant à la réalisation d'un comportement inné de la reproduction.
On observe également à la puberté, à partir de l'âge de 12-13 ans, lors de la sécrétion des hormones sexuelles par les gonades (gonadarche), une augmentation de l'activité sexuelle. Cet effet dépendrait principalement, surtout chez l'homme, de la testostérone. Néanmoins, l'expérimentation animale a montré que cet effet n'est pas spécifique d'un comportement de reproduction, car la suppression de la testostérone supprime tant les activités hétérosexuelles qu'homosexuelles (ARON 1996). De plus, la suppression clinique ou pathologique de cette hormone chez l'homme diminue fortement et parfois supprime le désir sexuel (BAGATELL & al. 1994; GIULIANO & al. 2004). Ces données suggèrent que l'effet de la testostérone est plutôt de type motivationnel (BANCROFT 2005), général, et ne concerne pas les séquences motrices spécifiques permettant d'aboutir au coït vaginal fécondant.
Par ailleurs, comme on l'a déjà vu, on observe que chez les chimpanzés pan paniscus (Bonobo) près d'un quart des activités sexuelles sont réalisées par des individus prépubères (HASHIMOTO 1997 ; DE WAAL 1990). Ensuite – et surtout – chez l'Homme, dans certains groupes sociaux sexuellement libéraux, les enfants ont des activités sexuelles dès l'âge de 3-4 ans. (SUGGS 1966 ; MALINOWSKI 1970 ; HENRY & HENRY 1974 ; KINSEY & al. 1948 ; MARTINSON 1994) (données détaillées à la section "1.a – Description des comportements de reproduction", page 3). Ces données montrent sans ambiguïté que les phénomènes hormonaux de la puberté, bien qu'ils augmentent significativement l'intensité du comportement érotique, ne sont pas à son origine.
En conclusion, contrairement à ce que suggère une simple observation du comportement des adolescents des sociétés occidentales, la puberté, phase de croissance de l'organisme et de maturation de l'appareil reproducteur (THIBAULT & LEVASSEUR 1991, 2001), ne serait pas une phase cruciale du comportement de reproduction. Chez l'Homme, les fonctions des hormones sexuelles de la puberté seraient essentiellement la maturation des organes reproducteurs, la maturation de la physiologie de la reproduction, et la modulation de certains aspects motivationnels du comportement sexuel.
Rôle central ou modulateur des hormones sexuelles
Une question cruciale par rapport au rôle des hormones sexuelles dans les aspects comportementaux de la reproduction est de savoir si les effets comportementaux observés dépendent uniquement et directement de ces hormones, ou bien s'ils dépendent d'un autre phénomène, et les hormones sexuelles ne seraient que des hormones modulatrices ou ayant un rôle secondaire qui reste à préciser.
Par exemple, par rapport à l'effet apparemment motivationnel de la testostérone, on pourrait envisager deux possibilités :
Première possibilité : les éléments biologiques à l'origine de la motivation sexuelle sont spécifiquement organisés autour de la testostérone. La motivation sexuelle dépend principalement de la testostérone. La testostérone joue un rôle clé dans l'initiation et la dynamique de la motivation sexuelle. La motivation sexuelle ne peut exister que lorsque, par exemple, le niveau de testostérone atteint un certain seuil, à l'adolescence. Exprimé autrement, un ensemble de facteurs innés concourt à l'apparition génétiquement programmée d'une motivation à la reproduction qui ne s'exprimera qu'à la puberté, grâce à la testostérone, et cela quelles que soient les caractéristiques de l'environnement.
Seconde possibilité : les éléments biologiques à l'origine de la motivation sexuelle ne sont pas spécifiquement organisés autour de la testostérone. La testostérone n'a qu'un rôle modulateur ou secondaire. La motivation sexuelle ne dépend pas principalement de la testostérone. Les fluctuations du niveau de testostérone ne peuvent qu'augmenter ou diminuer l'intensité de la motivation sexuelle, mais ne pourront ni créé ni supprimer cette motivation. La motivation sexuelle se développera principalement en fonction des caractéristiques de ses facteurs spécifiques, vraisemblablement la stimulation des zones érogènes qui active les processus de renforcement. Exprimé autrement, l'existence, l'initiation, le développement et la dynamique de la motivation érotique dépendent principalement de facteurs autres que les hormones sexuelles. Les fluctuations au cours de la vie du niveau de la testostérone ne pourront que moduler, uniquement si elle existe, l'intensité d'une motivation érotique acquise.
On peut objecter à cette seconde possibilité que la suppression expérimentale de testostérone diminue et souvent supprime la motivation sexuelle, suggérant que la molécule de testostérone est essentielle à l'activité de la structure neurale sous-tendant le processus de la motivation sexuelle. Mais, comme il existe toujours un niveau minimal de testostérone dans l'organisme, cette particularité expérimentale ou pathologique n'existe pas en situation naturelle. Ces expériences prouvent que la testostérone intervient dans la motivation sexuelle, mais ne donnent pas d'indications sur le rôle exact, primordial ou seulement modulateur, de cette hormone.
De plus, il a été montré expérimentalement que l'action des androgènes sur la libération de dopamine dans l'aire préoptique médiale semble faire intervenir le monoxyde d'azote (NO) par l'intermédiaire d'une probable androgénodépendance de son enzyme de synthèse, la NO synthase (DU & HULL 1999 ; GIULIANO & al. 2004). Ce résultat corrobore fortement l'hypothèse d'un rôle modulateur des androgènes. Par ailleurs, on observe que la copulation et l'éjaculation des mâles de la souche de souris B6D2F1 sont indépendantes des androgènes (OLSEN 1992), ce qui corroborent davantage le rôle modulateur, et non primordial, des hormones sexuelles. En effet, si le contrôle neural de la copulation était organisé autour de la testostérone, comment des différences aussi minimes que celles existant entre des souches de souris peuvent-elles être à l'origine d'une nouvelle organisation neurale fonctionnelle indépendante des androgènes ? Si la testostérone n'a qu'un rôle modulateur, il est possible de concevoir une hypothèse plausible qui rend compte de ce fait expérimental : par exemple, en se basant sur les résultats de Du & Hull (1999) présenté juste ci-dessus, une simple mutation sur l'enzyme NO synthase, qui rendrait cette dernière indépendante de l'action modulatrice des androgènes.
Ces données, et d'autres non présentées, montrent que les hormones sexuelles ont plutôt un rôle modulateur sur le comportement de reproduction, en modifiant la concentration et/ou l'activité des neurotransmetteurs et de la plupart des neuropeptides (BALTHAZART & FABRE-NYS 2001).
Conclusion
Les régulations hormonales, qui permettent de synchroniser les différentes composantes de la reproduction, entre-elles et avec les caractéristiques de l'environnement, existent toujours chez l'être humain. Néanmoins, au niveau comportemental, leurs effets résiduels sont relativement faibles, ne sont pas primordiaux, et, surtout, ne sont pas spécifiques du coït vaginal. Chez l'Homme, il semblerait que l'effet comportemental des hormones sexuelles – et en particulier la testostérone – soit surtout un effet de type motivationnel, général, qui tend à augmenter la fréquence toutes les activités sexuelles pratiquées par le sujet.
En conclusion, en fonction des connaissances actuelles, on ne connaît chez l'Homme aucun effet des hormones hypothalamo-hypophysaires ou épiphysaires qui permet un contrôle hormonal inné d'un ensemble organisé de séquences motrices destinées spécifiquement à la réalisation du coït vaginal reproducteur.
e – Processus émotionnels
À un niveau d'organisation encore supérieur du système nerveux, apparaissent des processus dits "émotionnels", caractérisés par des tendances psychocomportementales, accompagnées chez les humains par des sentiments affectifs, et générées par un nombre limité de systèmes neuraux ( Figure 1, niveau [6], page 14 ) (PANKSEPP 1998 ; DE BONIS 1996 ; HOLSTEGE & al. 1996 ; COSNIER 1994 ; DAMASIO 1995).
S'il existe un instinct de la reproduction, on pourrait éventuellement trouver, au niveau émotionnel, des structures neurales limbiques organisées de telle manière qu'elles peuvent initier des tendances comportementales globales permettant le coït vaginal. Une organisation spécifique pourrait être trouvée, par exemple, au niveau des structures impliquées dans la motivation.
En effet, afin que le comportement de reproduction puisse s'exécuter correctement, on peut supposer l'existence d'une motivation psychique innée (phénomène cérébral conscient) ou d'un "drive" comportemental inné (phénomène sensorimoteur inconscient) qui incite le sujet à effectuer le coït vaginal reproducteur. Cette motivation instinctuelle reproductrice doit soit, au maximum, inciter les partenaires hétérosexuels à effectuer le coït vaginal, soit, au minimum, les inciter à entrer en contact physique approprié (le coït serait alors ensuite provoqué par des réflexes déclenchés par le contact physique). Cette motivation pourrait être induite soit par des signaux spécifiques externes (synchronisateurs environnementaux), soit par des signaux ou des états spécifiques internes (tel l'œstrus et le rut chez certains animaux).
Or nous avons déjà montré plus haut qu'il n'existe pas, chez l'Homme, de stimuli-signaux spécifiques innés externes qui initient le comportement de reproduction hétérosexuel, qu'il n'existe pas de réflexes moteurs innés permettant le positionnement adéquat et l'intromission vaginale, et qu'enfin il n'existe pas non plus de connaissances innées relatives à la reproduction (voir la section suivante). La seule possibilité restante d’une motivation instinctuelle reproductrice serait alors l'existence innée d'une motivation ou d'un état interne induisant le rapprochement des partenaires hétérosexuels. La motivation instinctuelle reproductrice correspondrait alors à une motivation au contact hétérosexuel.
Exprimé autrement, la partie initiale du comportement de reproduction, c'est-à-dire le rapprochement sexuel de deux partenaires et l'initiation des activités sexuelles, pourrait être contrôlée par les hormones pubertaires par l'intermédiaire d'une motivation à rechercher une proximité physique ou un contact corporel avec un partenaire du sexe opposé. Le rôle des hormones sexuelles seraient limités à la phase appétitive du comportement sexuel. Les séquences suivantes du comportement de reproduction, c'est-à-dire la phase consommatoire, ne dépendraient plus des hormones sexuelles, mais d'autres processus, tels par exemple des réflexes déclenchés par le contact physique.
Or justement, d'après les données expérimentales présentées dans la section précédente "d – Régulation hormonale", nous avons vu que la testostérone est apparemment à l'origine d'une motivation sexuelle. Comme la testostérone contrôle également les phéromones, et que certaines phéromones voméronasales permettent de discriminer le partenaire de sexe opposé (DULAC & TORELLO 2003 ; STOWERS & al. 2002), la combinaison d'une motivation sexuelle avec des informations phéromonales peut finalement être à l'origine d'un comportement sexuel hétérosexuel.
Néanmoins, il existe deux problèmes qui semblent réfuter chez l'Homme l'hypothèse d'une motivation sexuelle innée, spécifique d'un comportement de reproduction. Le premier problème est la détérioration fonctionnelle du système voméronasal chez l'Homme (ZHANG & WEBB 2003), qui devrait le rendre incapable de discriminer de manière innée le partenaire reproducteur approprié. Le second problème, majeur, est la nature innée de cette motivation sexuelle. En effet, comment l'organisme humain (et également celui des autres mammifères) génère-t-il une motivation pour des activités motrices de stimulation génito-génitale avec un partenaire ? Même chez les mammifères inférieurs, avec une motivation hormonale innée, des phéromones innées qui discriminent le partenaire adéquat, plusieurs réflexes innés (lordose, érection, poussées pelviennes, éjaculation et d'ovulation réflexe), il manque un élément crucial qui est celui des actes moteurs permettant le contact physique génito-génital approprié. Or justement, nous avons vu (cf. "d – Le comportement de reproduction est-il inné", page 13) que c'est cette séquence de monte de la femelle par le mâle qui n'est pas innée et qui est apprise au cours du développement (WARD 1992). C'est également cette séquence qui est la plus difficile à préprogrammer en terme d'organisation neurale inné, car elle implique l'existence innée d'éléments cognitifs hautement complexes : "schéma corporel", "images préformées" ou connaissances spécifiques. Or nous avons déjà vu, et nous verrons encore dans la section suivante, que l'existence innée de ce type d'éléments cognitifs, en raison de leur inhérente complexité, est vraisemblablement improbable. En fonction de toutes ces données, une hypothèse plausible serait, chez tous les mammifères, que cette motivation sexuelle induite par la testostérone corresponde vraisemblablement à une motivation à ré-exécuter certaines séquences motrices de contacts corporels. Ces séquences auraient vraisemblablement eu lieu et auraient été apprises et renforcées principalement au cours de la période du développement.
En conclusion, dans l'état actuel des connaissances, on ne connaît pas chez l'Homme de motivation psychique, ou de drive moteur, innés, spécifiques de la reproduction, qui provoqueraient le coït vaginal reproducteur.
z – Processus cognitifs
Au niveau d'organisation le plus supérieur du système nerveux, apparaissent des processus dits "cognitifs", permettant des traitement élaborée de l'information (abstraction, catégorisation, planification, symbolisation...) ( Figure 1, niveau [7] et [8], page 14 ).
S'il existe un instinct de la reproduction, on devrait pouvoir trouver, à ce niveau cognitif, des réseaux neuronaux organisés de telle manière qu'ils contiennent des informations ou des connaissances spécifiques permettant de guider le comportement de reproduction vers le coït vaginal.
z.1 – Informations préformées
Existe-t-il à un premier niveau cognitif ( Figure 1, niveau [7] ) des informations innées et spécifiques, telles par exemples des images visuelles prototypiques, préformées dans les régions associatives sensorielles, et qui participent au contrôle du comportement de reproduction ?
En l'état des connaissances actuelles en neurosciences, cela semble difficilement plausible. Voir la section "1.d.a.2 – Vision", page 19, pour des explications détaillées concernant les images visuelles préformées [2]. Ces explications seraient également valables pour tous les autres types d'information préformées.
Chez les ovidés, les informations visuelles jouent également un rôle important dans l'attraction sexuelle, en particulier pour les mâles. Néanmoins, cette caractéristique semble acquise, dans la mesure où des animaux élevés dès la naissance par une mère d'une autre espèce (chevreau par brebis et agneau par chèvre) manifestent une attraction sexuelle pour des animaux de leur espèce adoptive et pas pour ceux de leur espèce génétique (KENDRICK & al. 1998). Ces données démontrent qu'il n'existe apparemment pas d'information innée spécifique du comportement de reproduction chez les ovidés.
Chez les primates non humains, les expériences classiques d'Harlow montrent que des primates élevés sans contacts avec leurs congénères présentent des réactions de peur en face du congénère et ne savent pas interagir avec les autres individus, tant au niveau social que sexuel (HARLOW 1958 ; 1959 ; 1970). Des singes rhésus élevés avec un substitut maternel en tissu interagissent socialement et sexuellement avec un substitut et non des congénères réels (DEUTSCH & LARSSON 1974). Ces données suggèrent que non seulement les primates doivent apprendre à discriminer le partenaire sexuel approprié, mais doivent en plus apprendre à reconnaître leurs congénères et apprendre à interagir avec eux. Il ne semble donc pas exister chez les primates non humains, et donc vraisemblablement chez l'Homme, d'information innée sociale ou sexuelle.
En conclusion, dans l'état actuel des connaissances, il ne semble pas pouvoir exister, chez l'Homme, d'informations cognitives préformées, innées, qui participeraient au contrôle du coït vaginal reproducteur.
z.2 – Connaissances innées
En tenant compte du développement particulièrement important chez l'Homme des capacités cognitives, il pourrait exister de manière innée des "connaissances" ou des "schémas mentaux" ( Figure 1, partie [8], page 14 ), au moins partiels, qui permettraient d'induire le coït vaginal.
Mais aucune donnée ne permet de supporter cette hypothèse. Les enquêtes réalisées auprès d'enfants et d'adolescents (VOLBERT 2000 ; voir également MARTINSON 1994 ; CONSTANTINE & MARTINSON 1981) montrent que les connaissances relatives à la reproduction doivent être apprises. Les données ethnologiques provenant de certaines sociétés traditionnelles pré-industrielles montrent que, non seulement l'Homme n'a aucune connaissance innée relative aux comportements nécessaires à la reproduction et au coït vaginal, mais que, de surcroît, il n'a aucune connaissance innée de la physiologie de la procréation sexuée et de la gestation. Dans ces sociétés, il existe diverses croyances erronées relatives à la procréation, comme par exemple que l'enfant provient de l'incarnation d'un esprit dans le ventre de la femme.
En conclusion, dans l'état actuel des connaissances, il ne semble pas pouvoir exister, chez l'Homme, de connaissances préformées, innées, qui participeraient au contrôle du coït vaginal reproducteur.
h – Archétype du comportement de reproduction mammalien
Les analyses précédentes ont montré qu'il n'existait pas, même chez les mammifères non humains, une organisation neurale qui permet la réalisation préprogrammé d'un comportement de reproduction. Néanmoins, surtout chez les mammifères les plus simples, on observe qu'il existe de nombreux éléments innés, tant instinctuels que physiologiques ou autonomes, qui peuvent participer directement et indirectement à la réalisation appropriée du coït vaginal fécondant : l'inhibition saisonnière par la mélatonine, les hormones sexuelles, les phéromones sexuelles, la lordose, l'érection, les poussées pelviennes, le réflexe éjaculatoire, l'ovulation réflexe lors du coït ...
À partir de ces différents éléments, chacun relativement simple, mais nombreux et complémentaires, il semble apparaître un véritable comportement de reproduction, dont le seul but et les seules séquences comportementales sont la réalisation d'une copulation hétérosexuelle fécondante, effectuée à la saison propice. Néanmoins, il ne semble pas exister d'instinct, c'est-à-dire une programmation centrale innée des différentes séquences motrices nécessaires au comportement de reproduction. Il existerait plutôt une possibilité de coordination de différents modules fonctionnels, innés mais élémentaires, qui permet, après quelques apprentissages indispensables, l'émergence d'un comportement de reproduction.
À partir de toutes les données recueillies dans les études précédantes, il semble possible de modéliser les facteurs et la dynamique de l'archétype du comportement de reproduction des mammifères.
Facteurs primordiaux inné du comportement de reproduction
Quels seraient les facteurs primordiaux à l'origine du comportement de reproduction et quel seraient leurs fonctions respectives ?
Les analyses effectuées dans les sections précédentes ont mis en évidence l'existence de 5 facteurs primordiaux innés : les hormones, les phéromones, les réflexes sexuels, les interconnexions précablées et les processus de renforcement.
– Les hormones. Elles régulent et coordonnent la plupart des autres facteurs du comportement de reproduction (SIGNORET & BALTHAZART 2001, 1991). Elles régulent principalement :
– L'activation du comportement sexuel durant :
– la saison propice à la reproduction (ALEANDRI & al. 1996).
– La période favorable du cycle physiologique de l'appareil reproducteur (période œstrale).
– Une motivation sexuelle générale (mais qui serait acquise).
– La facilitation des réflexes sexuels.
– L'émission des phéromones sexuelles.
– Les phéromones. Elles permettraient surtout le déclenchement de la motivation sexuelle (via le système olfactif principal et voméronasal ; MONCHO-BOGANI & al. 2004 ; YOON & al. 2005) et la discrimination d'un partenaire reproducteur adéquat (via le système voméronasal ; STOWERS & al. 2002 ; DULAC & TORELLO 2003).
– Les réflexes sexuels (moteurs, autonomes et neuroendocriniens).
Les réflexes sexuels moteurs permettent la réalisation de la dernière phase de la copulation. Ces réflexes sont :
– La lordose (PFAFF & al. 1994).
– L'immobilisation (de la femelle).
– L'intromission (MEISEL & SACHS 1994).
– Les poussées pelviennes.
Les réflexes sexuels autonomes et neuroendocriniens facilitent la copulation et permettent la fécondation. Ces réflexes sont :
– L'érection et la lubrification (GIULIANO & RAMPIN 2004).
– L'éjaculation (COOLEN 2005).
– L'ovulation réflexe, provoquée par le coït (SPIES & al. 1997).
– Des interconnexions précablées. Elles permettent de relier entre elles les différentes structures neurales impliquées dans le comportement de reproduction. Apparemment, comme pour tous les précablages, seules les connexions globales seraient innées (CHILTON 2006). Les connexions précises et fonctionnelles seraient acquises, vraisemblablement par un mécanisme semblable à celui de la stabilisation sélective (CHANGEUX & DANCHIN 1976).
– Les processus de renforcement. Ces processus, associés à la peau poilue (OLAUSSON & al. 2002), aux zones érogènes, à la copulation (CAGGIULA & HOEBEL 1966) ainsi qu'à l'éjaculation et à l'orgasme (HOLSTEGE & al. 2003), seraient à l'origine de la réitération de toutes les activités de stimulation du corps et surtout des zones génitales. Ces processus seraient également, mais indirectement, à l'origine de l'apprentissage de la plupart des séquences non innées du comportement de reproduction.
Facteurs primordiaux acquis du comportement de reproduction
Par ailleurs, les analyses effectuées dans les sections précédentes ont mis en évidence des éléments cruciaux du comportement de reproduction qui ne seraient pas innés. Ces éléments, qui seraient acquis au cours de la période du développement de l'organisme, seraient :
– La reconnaissance des congénères (KENDRICK & al. 1998).
– La motivation sexuelle à rechercher le contact physique génital.
– La socialisation sexuelle (SPEVAK & al. 1973).
– La capacité au positionnement génito-génital approprié des corps (GRUENDEL & ARNOLD 1969 ; MISSIAKAN 1969 ; TURNER & al. 1969).
En fonction des connaissances et de la compréhension actuelle du fonctionnement du système nerveux, il est vraisemblable et cohérent que ces capacités doivent être acquises. En effet, on remarque que ces capacités sont en fait celles qui seraient les plus difficiles à précabler ou à coder génétiquement. Ces capacités nécessiteraient l'existence innée de connaissances ou de représentations élaborées du corps et du partenaire, qui ne pourraient être codées que dans les régions les plus complexes du système nerveux. En fonction des connaissances neurobiologiques actuelles, l'implémentation innée de telles capacités cognitives aussi complexes est difficilement explicable. (cf. la section "z – Processus cognitifs", page 36).
Par contre, il est très probable que ces capacités cognitives seraient acquises grâce aux diverses et très fréquentes interactions corporelles, sociales et sexuelles avec les congénères (mouvements et contacts physiques permanents avec les autres nouveau-nés, léchage ano-génital par la mère (MOORE 1992), investigation régulière phéromone-dépendante de la région anogénitale des congénères (SPEVAK & al. 1973), jeux corporels (HARD & LARSSON 1971), jeux sexuels ...), ayant lieu durant toute la période du développement (WARD 1992 ; VANDERSHUREN & al. 1997). De plus, l'apprentissage de ces capacités induirait, au moins en partie, le développement des structures neurales impliquées dans le contrôle du comportement sexuel (amygdale médiale, noyau préoptique médian, noyaux moteurs médullaires ... ; COOKE & al. 2000 ; MOORE & al. 1992 ; BAUM & al. 1995).
De cette manière, l'interaction entre l'environnement et les facteurs innés permet le développement structurel et fonctionnel du système nerveux, et, à la puberté, tous les facteurs et processus nécessaires à la réalisation du comportement de reproduction seraient fonctionnels.
Émergence du comportement de reproduction
La première phase de l'émergence du comportement de reproduction se déroulerait durant la période du développement de l'organisme. C'est durant cette phase que les éléments non innés du comportement de reproduction seraient acquis.
La motivation sexuelle au contact physique et génital serait acquise grâce au fait que la stimulation corporelle, en particulier au niveau des zones génitales, serait particulièrement renforçante. Les contacts corporels postnataux avec la mère et avec les autres nouveau-nés, et en particulier le léchage ano-génital (MOORE 1992 ; BAUM & al. 1995 ; WARD 1992) ainsi que les jeux sexuels avec les autres congénères, développeraient graduellement une motivation sexuelle à rechercher des stimulations génitales. Cette motivation sexuelle acquise serait ensuite fortement potentialisée à la puberté sous l'effet des hormones sexuelles, vraisemblablement par une action modulatrice sur les enzymes contrôlant la synthèse de certains neuromédiateurs (DU & HULL 1999).
La socialisation sexuelle, c'est-à-dire l'ensemble des attitudes et des réactions appropriées à l'interaction sociale sexuelle avec les congénères, serait apprise au cours des nombreux jeux sociaux (VANDERSCHUREN & al. 1997). Ces activités ludiques sont fréquemment répétées car elles sont renforçantes, en particulier durant la période du développement (DOUGLAS & al. 2004).
Le positionnement génito-génital (la séquence de la monte chez les mammifères inférieurs) serait appris au cours des interactions corporelles et des jeux sexuels avec les congénères. Ces activités seraient fréquemment répétées en raison de leur caractère renforçant (DOUGLAS & al. 2004). Les premiers contacts corporels post-nataux et plus particulièrement tous les types de contacts et d'explorations corporelles effectués au cours des jeux sociaux pourraient être à l'origine de la construction d'un "schéma corporel" [3] de l'individu et des congénères. L'apprentissage d'un schéma corporel permettrait à l'animal de pouvoir réaliser des ajustements posturaux adaptés lors d'interactions corporelles spécifiques avec ses congénères (toilettage, agression, copulation ...). Au cours de ces interactions corporelles, toutes celles en relation avec les régions génitales ou des réflexes sexuels pourraient amorcer des séquences sexuelles partielles (jeux sexuels), qui seraient progressivement intégrées dans des schèmes moteurs plus globaux (HARD & LARSSON 1971). La motivation sexuelle pour la région génitale pourrait provenir en partie des fréquentes explorations, provoquées par des phéromones, de la région anogénitale. Quant à l'intromission effectuée juste après la monte, elle serait rendu possible, entre autres, par des stimulations postnatales ayant permis le développement de noyaux médullaires moteurs (MOORE & al. 1992).
Ainsi, les éléments non innés du comportement de reproduction serait appris au cours du développement, et, à la puberté, tous les facteurs et processus nécessaires à la réalisation de ce comportement seraient fonctionnels.
Chez les mammifères inférieurs, à la puberté, qui est la seconde phase de l'émergence du comportement de reproduction, la dynamique archétypique du comportement sexuel serait la suivante :
Les hormones auraient principalement un rôle, par rapport au comportement, de coordination et de modulation, en rendant actif de manière simultané les différents processus neurobiologiques impliqués dans le comportement de reproduction. La mélatonine, par son action sur les neurones à GnRH, activerait le comportement sexuel durant la saison favorable à la reproduction (ALEANDRI & al. 1996). Les hormones sexuelles lèveraient les inhibitions toniques sur les réflexes sexuels, abaisseraient les seuils sensoriels, déclencheraient la synthèse et la libération des phéromones sexuelles et potentialiseraient la motivation sexuelle (KNOBIL & NEILL 1994). L'organisme est ainsi prêt pour effectuer les séquences comportementales aboutissant à la fécondation.
Puis, lorsque les animaux sexuellement naïfs seraient en présence, les phéromones seraient les principaux signaux primaires qui permettraient le déclenchement de la motivation sexuelle innée et acquise (MONCHO-BOGANI & al. 2005, 2004, 2002) et la discrimination du partenaire sexuel approprié (STOWERS & al. 2002 ; DULAC & TORELLO 2003).
Enfin, lorsque les animaux seraient en contact physique, les stimuli corporels induiraient la monte et le déclenchement successifs des différents réflexes sexuels : lordose (PFAFF & al. 1994), intromission, poussées pelviennes, éjaculation (ALLARD & al. 2005) et libération de l'ovule (SPIES & al. 1997).
On observe ainsi que le comportement de reproduction émergerait principalement de la coordination fonctionnelle de différents réflexes et processus sexuels innés, d'interactions avec les congénères et de différents apprentissages qui sont toujours réalisés dans l'environnement écologique normal.
Remarques complémentaires
Pour un observateur extérieur, le comportement de reproduction apparaîtrait comme inné, dans la mesure où les apprentissages, tels ceux induits par le léchage ano-génital, ne sont pas apparents, ne sont pas directement liés à la reproduction ou ne sont pas considérés comme "sexuels".
Suivant les espèces, on observe que certains éléments, telle l'ovulation réflexe après le coït, sont absents. Néanmoins, tant que les éléments absents sont mineurs, le comportement n'est guère affecté et la reproduction reste toujours possible.
Une fois que l'animal naïf à réalisé un premier comportement sexuel, et au fur et à mesure de l'accroissement de son expérience sexuelle et des apprentissages concomitants, les différentes séquences motrices seront exécutées plus efficacement, d'autre signaux (visuels, auditifs, gustatifs, etc.) pourront devenir sexuels par conditionnement, et le comportement de reproduction pourra être effectué malgré l'absence de certains signaux cruciaux pour l'animal sexuellement naïf.
Concept d' "Instinct partiel"
L'étude ci-dessus des facteurs innés et acquis à l'origine du comportement de reproduction suggère qu'il n'existerait pas d'instinct, c'est-à-dire une programmation centrale innée des différentes séquences motrices nécessaires à la réalisation de ce comportement. Il existerait plutôt une possibilité de coordination de différents modules fonctionnels, innés mais élémentaires, qui permet, après quelques apprentissages indispensables, l'émergence d'un comportement de reproduction.
Ces caractéristiques particulières nous amènent à proposer le concept d'un "instinct partiel", c'est-à-dire un ensemble incomplet d'éléments innés, mais, parce qu'ils sont associées à des circonstances environnementales spécifiques qui existent quasiment toujours au cours du développement (par exemple les contacts corporels, les stimuli sensoriels, les interactions mère-enfant ...), les éléments manquant sont indirectement toujours acquis et le comportement "partiellement instinctuel" s'exécute correctement à sa période de maturité, sans nécessité d'apprentissages spécifiques.
Évolution du comportement de reproduction & Extrapolation à l'Homme
En étudiant les modifications de l'organisation du système nerveux des différentes espèces de mammifères au cours de l'évolution, on remarque d'importants changements structurels et fonctionnels qui influencent directement les facteurs primordiaux du comportement de reproduction.
Les principales modifications, des rongeurs à l'Homme, sont les suivantes :
– Un important développement du prosencéphale.
– Une diminution importante de l'influence des hormones et des phéromones.
– Une détérioration fonctionnelle du système voméronasal.
L'important développement du néocortex, siège des fonctions cognitives, induit une augmentation de l'importance des facteurs cognitifs dans le comportement de reproduction. Apparemment, plus le néocortex est développé dans une espèce donnée, plus on observe de variations et d'adaptations comportementales dues aux capacités d'analyse du contexte et de prise en compte de l'expérience individuelle. Chez l'Homme, le cortex, qui s'est développé jusqu'à constituer les trois quarts du cerveau, serait le support ayant permis l'émergence de la culture et de son influence déterminante dans la sexualité humaine.
Quant aux facteurs hormonaux, qui jouent un rôle majeur dans le contrôle du comportement de reproduction, on remarque que leur influence décroît graduellement jusqu'à devenir mineure chez l'être humain. Le contrôle saisonnier a quasiment disparu : l'activité sexuelle et la reproduction humaine existent tout au long de l'année et on n'observe, seulement dans les pays nordiques, qu'un effet mineur des saisons sur le comportement de reproduction (ALEANDRI & al. 1996 ; PANDI-PERUMAL & al. 2006). Le contrôle œstral est affaibli : la femme peut avoir des activités sexuelles tout au long de son cycle, même si on observe encore un effet hormonal dans la période de fécondité (WILCOX & al. 2004). Les hormones pubertaires ne sont plus déterminantes dans l'initiation du comportement sexuel : tant chez les chimpanzés (HASHIMOTO 1997 ; DE WAAL 1990) que chez l'Homme (SUGGS 1966 ; MALINOWSKI 1970 ; MARTINSON 1994), si le contexte le permet, les activités sexuelles débutent dès les premières années de la vie. Enfin, le réflexe neuroendocrinien d'ovulation provoqué par le coït n'est plus fonctionnel chez de nombreuses espèces de mammifères, dont les primates.
L'altération du système voméronasal chez les primates de l'Ancien Monde (ZHANG & WEBB 2003) provoque la dysfonction ou la disparition des fonctions assurées par cet organe, en particulier la discrimination du partenaire reproducteur approprié (STOWERS & al. 2002 ; DULAC & TORELLO 2003). De plus, bien que des phéromones puissent encore être perçues au niveau du système olfactif principal (LIBERLES & BUCK 2006), on observe que leurs effets, en particulier comportementaux, sont faibles chez l'être humain (FOIDART & al. 1994).
Il ne reste donc, chez l'Homme, que les facteurs primordiaux innés à l'origine de la dernière partie du comportement de reproduction : les réflexes sexuels (érection, lubrification, intromission, poussées pelviennes, éjaculation), qui permettent la réalisation de la copulation. Et il reste également les processus de renforcement, associés à la peau poilue et aux zones érogènes, qui sont à l'origine de la réitération des activités de stimulation du corps et surtout des zones génitales.
Quelles règles générales concernant l'évolution du comportement de reproduction des mammifères peut-on déduire de toutes ces observations ?
Il apparaît que les différents facteurs primordiaux sont relativement indépendants les uns des autres et peuvent être modifiés ou disparaître, sans obligatoirement compromettre la reproduction. Dans ces cas, on observe que le comportement de reproduction se développe en fonction des caractéristiques des facteurs qui sont encore existants. Si le contrôle saisonnier s'affaiblit, le comportement s'exprime en continu mais avec des variations saisonnières de fréquence ; si la discrimination du partenaire approprié disparaît, le comportement devient bisexuel (DULAC & TORELLO 2003) ; etc. Apparemment, quelles que soient les modifications provoquées par l'évolution dans une espèce, tant qu'un comportement aboutissant au coït vaginal fécondant peut se développer à partir des caractéristiques modifiées des facteurs primordiaux, cette espèce peut survivre et se reproduire.
En conclusion, l'évolution du comportement de reproduction mammalien dépendrait des modifications ou des altérations que l'évolution de la structure du système nerveux provoque sur les différents facteurs primordiaux. Pour chaque espèce, les caractéristiques basiques du comportement de reproduction dépendraient des caractéristiques des facteurs primordiaux qui existent encore dans cette espèce.
Quelles caractéristiques du comportement de reproduction des primates peut-on déduire de ces tendances de l'évolution ?
En synthèse, chez les primates et surtout chez l'Homme, les facteurs primordiaux hormonaux et phéromonaux deviendraient secondaires, la séquence copulatoire serait conservée, et ce serait des facteurs cognitifs et de renforcement qui seraient à l'origine de la partie initiale (motivation, orientation) du comportement de reproduction. Ces évolutions importantes des principaux facteurs comportementaux devraient avoir un impact majeur sur le comportement de reproduction humain.
q – Conclusion
En conclusion, chez les mammifères inférieurs, il semblerait que les réflexes et les processus sexuels innés, la régulation et la coordination de ces éléments, ainsi que des apprentissages quasi systématiquement réalisés au cours du développement, permettraient, indirectement, le développement d'un comportement de reproduction qui apparaît comme étant inné.
Chez l'Homme, en raison des modifications évolutives, la dynamique à l'origine du comportement de reproduction semble avoir été profondément modifiée. En synthèse des données précédemment analysées, il semblerait que les effets cycliques, saisonniers et phéromonaux ne seraient plus que marginaux. Les hormones sexuelles n'agiraient plus que sur les aspects anatomiques et physiologiques des tissus et des organes liés à la reproduction, et elles n'interviendraient, au niveau comportemental, que dans la modulation de l'intensité de la motivation sexuelle acquise. Aucune donnée disponible ne corrobore l'hypothèse d'un contrôle hormonal ou instinctuel inné d'un ensemble organisé de séquences motrices destinées spécifiquement à la réalisation du coït vaginal reproducteur. Quant aux réflexes et aux processus sexuels innés encore existants, ils semblent insuffisants pour faire émerger, par l'intermédiaire de coordinations et d'acquisitions latentes ou indirectes, un comportement permettant la reproduction sans des apprentissages spécifiques.
Plus globalement, l'ensemble de toutes les données animales et humaines relatives à la reproduction montre qu'il est très probable que chez l'Homme, hormis le réflexe des poussées pelviennes chez le mâle, les différents aspects du comportement permettant la reproduction (en particulier les éléments les plus cruciaux : la motivation sexuelle, le rapprochement des partenaires et le coït vaginal) ne seraient ni instinctuels, ni même innés.
Chez l'Homme, la reproduction dépendrait d'une autre dynamique que celle mise en évidence par expérimentation chez les mammifères inférieurs, les oiseaux et les poissons.
L'ensemble des analyses et des données présentées dans ce chapitre réfuteraient toutes les théories innéistes relative au comportement de reproduction humain.

e – Apprentissage du comportement permettant la reproduction
Les données et les analyses précédentes ont montré que le comportement de reproduction n'était vraisemblablement pas inné chez l'Homme. Mais si ce comportement est acquis, comment s'apprend-il ? Quels sont les facteurs clés et la dynamique de cet apprentissage ?
Certaines théories sociologiques, telles celles de Ira Reiss, ou de John Gagnon et William Simon, supposent que l'essentiel – voire la totalité – du comportement sexuel humain est acquis (ALLGEIER & ALLGEIER 1992). Cependant ces théories sociologiques ne peuvent expliquer ni l'origine et la persistance dans toutes les sociétés humaines du plaisir érotique, de l'orgasme et des activités sexuelles génitales, ni l'évolution phylogénétique de la fonction des hormones et des phéromones sexuelles, ni les moyens biologiques absolument nécessaires à la réalisation de la fonction vitale et fondamentale de la reproduction chez les organismes sexués.
Il est donc nécessaire d'élaborer un nouveau modèle de l'apprentissage du comportement sexuel humain (WOODSON 2002), qui prenne en compte les facteurs biologiques cruciaux. Dans une approche pluridisciplinaire, l'analyse à la fois des données phylogénétiques, biologiques, du comportement sexuel, des connaissances en sexologie, des théories de l'apprentissage par conditionnement classique et opérant, ainsi que des théories de l'apprentissage social, permet, à partir d'éléments biologiques innés qui sont communs à tous les mammifères, de proposer un modèle de l'apprentissage du comportement permettant la reproduction.
Phylogenèse et extrapolation à l'Homme
Comme nous venons de le voir dans les chapitres précédants, on observe chez les mammifères que le comportement de reproduction dépend de la coordination et de l'interaction de plusieurs facteurs biologiques : hormones sexuelles, cycles physiologiques, phéromones sexuelles, réflexes élémentaires innés (lordose, érection, poussées pelviennes, …), et processus de renforcement.
Un certain nombre de données, déjà présentées dans la section "Évolution du comportement de reproduction", suggèrent que l'importance relative de ces différents facteurs change en raison de la modification de l'architecture du système nerveux au cours de l'évolution phylogénétique. Ces modifications neurales structurelles entraîneraient des modifications fonctionnelles. Chez l'Homme, en raison de l'importance du développement des structures prosencéphaliques, et tout particulièrement du néocortex, certains facteurs deviendraient mineurs (hormones, cycles physiologiques, olfaction, phéromones …), tandis que d'autres deviendraient prépondérants (processus de renforcement, facteurs hédoniques, processus cognitifs …) (Figure 21). Les modifications de l'importance relative de ces facteurs structuro-fonctionnels provoqueraient une modification de la dynamique et des caractéristiques du comportement permettant la reproduction. Les mêmes facteurs, mais avec une importance relative différente, induiraient dans chaque ordre de mammifère le développement de comportements sexuels sensiblement distincts les uns des autres. (Voir également les travaux d'Insel (1997) relatif à l'attachement, pour un autre exemple bien étudié de modifications structurelles qui entraînent d'importantes modifications fonctionnelles).
Hypothèse de l'apprentissage
En fonction de ces données phylogénétiques, où les facteurs de renforcement, hédoniques et cognitifs seraient devenus prépondérants chez l'Homme, quelle hypothèse peut-on proposer qui rendrait compte de l'apprentissage du comportement permettant la reproduction ?
Dans un premier temps, l'observation du comportement sexuel humain montre que la sexualité consiste essentiellement, au niveau le plus basique, en des stimulations hédoniques du corps : caresses sensuelles, baisers, activités oro-génitales, coït, etc. (ALLGEIER & ALLGEIER 1992 ; COMFORT 1992). Les études menées en sexologie dans différents groupes sociaux ou ethniques indiquent que, quelles que soient les variations des comportements sexuels, ils consistent toujours en la stimulation de certaines zones particulières du corps : les zones érogènes (FORD & BEACH 1965). Cette première analyse permet de caractériser le pattern basique du comportement sexuel humain, qui serait la répétition de la stimulation des zones érogènes.
Dans un second temps, il existe en neurobiologie et en psychologie des théories qui expliquent la répétition des actions. La théorie psychologique de la récompense (reward) et la théorie neurobiologique des renforcements, qui correspondent vraisemblablement au même phénomène cérébral, prédisent que tout comportement qui est renforcé ou récompensé sera répété. Cette seconde analyse permet de formuler l'hypothèse que la stimulation des zones érogènes procure une récompense qui incite le sujet à répéter cette stimulation. Cette hypothèse permet d'expliquer la répétition des stimulations des zones érogènes du corps, activité qui constitue l'aspect primordial de la sexualité humaine.
Un certain nombre de données corroborent cette hypothèse :
– Des preuves expérimentales de l'implication des processus de renforcement dans les comportements sexuels, chez tous les mammifères.
– L'importance fonctionnelle des zones érogènes, et surtout du pénis / clitoris dans la dynamique du comportement sexuel.
– L'importance de la relation fonctionnelle entre les zones érogènes et les processus de renforcement.
a – Processus de renforcement
Les renforcements et les conditionnements sont des processus majeurs dans la dynamique psychique et comportementale chez l'Homme. Ces processus interviennent dans l'acquisition de nombreux comportements, dont le comportement sexuel (CRAWFORD & al. 1993).
De nombreuses données expérimentales corroborent l'implication des processus de renforcement (niveau neurobiologique) et des récompenses (niveau psychique) dans le comportement sexuel (pour une revue, voir PFAUS & al. 2001). Ces renforcements et ces récompenses, activés par la stimulation des zones érogènes, sont appelés renforcements érotiques ou récompenses érotiques dans les textes suivants.
Pour donner quelques exemples, chez les rongeurs, en utilisant le protocole expérimental de préférence de place (place preference), on observe que la copulation est une récompense érotique pour la rate (OLDENBURGER & al. 1992 ; PAREDES & ALONSO 1997), et que la copulation jusqu'à l'éjaculation est, vraisemblablement par l'intermédiaire des opioïdes endogènes, une récompense érotique pour le rat mâle (AGMO & BERENFELD 1990 ; AGMO & GOMEZ 1993). De plus, l'intromission sans éjaculation produirait également une récompense érotique, car cette activité est suffisante pour inciter un rat à réaliser un apprentissage dans un labyrinthe (WHALEN 1961). Par ailleurs, la stimulation dans le faisceau médian du télencéphale au niveau de l'hypothalamus postérieur provoque une récompense érotique chez le rat (CAGGIULA 1970 ; CAGGIULA & HOEBEL 1966). Chez les primates, on observe que la section du nerf dorsal du pénis provoque chez le macaque rhésus une diminution de la fréquence des copulations, malgré la persistance de l'éjaculation. Ce résultat suggère que la copulation est une récompense érotique et que les afférences péniennes ont un effet plus renforçant que l'éjaculation (MEISEL & SACHS 1994 p. 30). Enfin, chez l'Homme, on observe grâce à l'imagerie cérébrale qu'au moment de l'éjaculation et de l'orgasme les régions impliquées dans les renforcements sont actives (HOLSTEGE & al. 2003).
Les renforcements érotiques sont extrêmement puissants déjà chez les mammifères les plus simples (CRAWFORD & al. 1993). Par exemple, uniquement en utilisant la copulation comme récompense chez des rats ou des lapins, il est possible de les conditionner expérimentalement à copuler avec d'autres espèces animales ou avec des objets (FORD & BEACH 1965, p. 152).
À noter par ailleurs que les processus de renforcement interviendraient également de manière indirecte dans l'apprentissage du comportement de reproduction, en rendant appétitif les jeux sociaux qui permettent l'apprentissage des interactions sociales sexuelles appropriées au contexte (VAN DEN BERG & al. 1999 ; DOUGLAS & al. 2004 ; VANDERSCHUREN & al. 1997).
De plus, il existe également des relations neuroanatomiques directes entre les processus de renforcement et les neurones de l'hypothalamus qui contrôlent le système hormonal de la reproduction (neurones à GnRH / LHRH – BOEHM & al. 2005). Ces données montrent que les processus de renforcement participent également à la physiologie de la reproduction.
Même si on ignore actuellement le fonctionnement exact de ces processus de renforcements, les nombreuses données éthologiques, psychologiques et expérimentales montrent qu'ils existent chez tous les mammifères et qu'ils participent de manière significative à la dynamique du comportement sexuel.
b – Zones érogènes
Les zones érogènes, qui sont des régions du corps spécifiques, et dont la stimulation est à l'origine des sensations érotiques, sont un fait bien établi en sexologie (ZWANG in BRENOT 2004).
L'importance des zones érogènes, et tout particulièrement des zones génitales, a été mis en évidence par les travaux de Masters & Johnson. L'observation et la mesure de différents paramètres anatomiques et physiologiques, au cours de plus de 10.000 cycles de réponse sexuelle auprès de 694 hommes et femmes, ont montré que le pénis et le clitoris étaient les principales régions sources du plaisir sexuel (MASTERS & JOHNSON 1980).
Par ailleurs, le pénis et le vagin sont bien adaptés au coït, grâce à leur conformation anatomique complémentaires (SCHULTZ 1999), à l'érection pénienne, à la lubrification vaginale, et à la position du clitoris qui est indirectement stimulé lors du coït (MASTERS & JOHNSON 1980).
Ces données bien établies permettent de prévoir que les stimulations érotiques du corps devraient principalement concerner les régions génitales et que le coït vaginal, absolument nécessaire à la reproduction de l'espèce, devrait être une des activités sexuelles les plus pratiquées.
g – Voies de liaison
Les voies centrales de projections des récepteurs des zones érogènes ne sont pas connues avec précision. Les études neuroanatomiques détaillées disponibles concernent les projections des nerfs commandant l'érection des corps caverneux du pénis (MARSON & al. 1993) et du clitoris (MARSON 1995) chez le rat, et rien ne permet dire que ces projections sont parallèles à celles des récepteurs des zones érogènes du pénis ou du clitoris.
Néanmoins, des données expérimentales chez les rongeurs (COOLEN 2005 ; MALLICK & al. 1994) confirment l'existence de projections fonctionnelles vers le diencéphale, et, surtout, des données cliniques humaines corroborent l'existence de relations anatomiques entre les zones érogènes des organes génitaux et des structures cérébrales en relation avec les processus de renforcement. Ces données humaines montrent que cette relation fonctionnelle est capitale pour l'expression du comportement sexuel.
– Dans les cas de section accidentelle de la moelle épinière au-dessus de la dixième vertèbre thoracique, les zones érogènes génitales et anales deviennent insensibles, laissant fonctionnelle uniquement la zone érogène buccale. Les données cliniques indiquent bien une disparition des sensations et des activités érotiques génitales, ainsi qu'une réorganisation du comportement érotique autour des zones érogènes orales. On observe également l'apparition de nouvelles zones érogènes (transfert érogène), autour desquels s'organise de nouvelles activités érotiques. Le coït vaginal est encore pratiqué, mais il est réalisé de manière volontaire, dans l'objectif de procréer ou de satisfaire un partenaire qui n'est pas handicapé. Le sujet lésé n'a plus de motivation à réaliser le coït (SOULIER 2001 ; SIPSKI 1998, 2001 ; SIPSKI & al. 2001).
– La pathologie du spina-bifida se caractérise principalement par une malformation vertébrale, entraînant l'écrasement de la moelle épinière entre des vertèbres ou par le liquide céphalo-rachidien. Dans certains cas l'atteinte neurologique provoque une absence de la sensibilité de la région génitale. À la différence des sections médullaires accidentelles, ces personnes atteintes n'ont jamais, au cours de leur vie, éprouvé de sensations génitales. On observe alors que les patients ne se masturbent pas et ne sont pas intéressés par ce type d'activité. De plus, l'orgasme génital est absent. Le coït vaginal, lorsque la pathologie n'a pas altéré les réflexes sexuels, n'est réalisé que de manière volontaire dans l'intention de procréer. On observe donc que l'activité érotique est absente des zones érogènes insensibles. Par contre, les patients perçoivent et recherchent des sensations de type érotique (‘frissons ou sensations particulières’ ; ‘impression de bouffées de chaleur’ ; para-orgasme) provoquées par la stimulation de la partie haute du corps (SOULIER 2001 ; LABAT & MAUDUYT DE LA GRÈVE 1996 ; CASS & al. 1986). On observe donc que l'activité érotique s'organise autour des zones érogènes préservées ou nouvelles.
Ces données cliniques suggèrent que des informations somatosensorielles cruciales pour le comportement permettant la reproduction sont véhiculées par des faisceaux ascendants de la moelle épinière (éventuellement les mêmes voies que celles de la sensibilité générale : le faisceau du lemnisque médian jusqu'au cortex sensoriel pour la sensibilité épicritique et le faisceau spinothalamique ventral pour la sensibilité protopathique). Ces données suggèrent également – et surtout – l'importance cruciale et primordiale des stimulations somatosensorielles dans le comportement sexuel humain.
d – Conclusion
L'ensemble de toutes les données relatives à la reproduction, présentées dans tous les chapitres précédents, suggère que chez l'Homme le comportement permettant le coït vaginal reproducteur ne serait pas inné, mais qu'il serait acquis, par un apprentissage de type instrumental, grâce principalement aux processus de renforcements. Ces processus, en relation anatomique avec les zones érogènes, seraient à l'origine du développement et de l'apprentissage d'un comportement de stimulation de ces zones, avec une préférence particulière pour le pénis / clitoris.
Cette hypothèse est présentée et explicitée plus en détail dans le chapitre suivant.
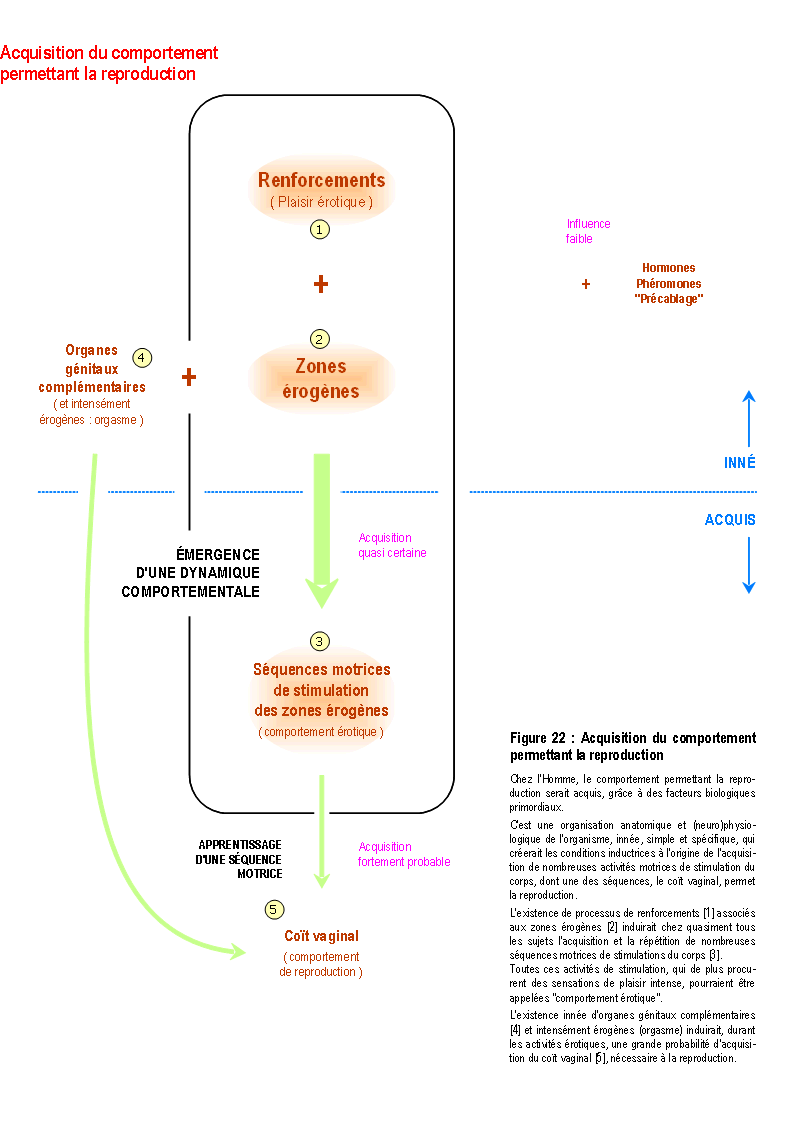
 |
Figure 22
2 – Présentation des hypothèses et du modèle comportemental
L'objectif de ce chapitre est de présenter l'hypothèse principale de ce travail de recherche. Comme cette hypothèse est nouvelle et que la dynamique comportementale qu'elle implique est particulière, la seconde partie de ce chapitre présente ce nouveau modèle de l'apprentissage du comportement permettant la reproduction. Cette explicitation complète, par la description d'étapes et de séquences concrètes, devrait permettre au lecteur de mieux comprendre l'hypothèse principale.
a – Présentation synoptique de l'hypothèse principale
L'hypothèse principale de la thèse, formulée de la manière la plus synthétique, est la suivante :
Chez l'Homme, l'acquisition du comportement permettant la reproduction dépend – principalement mais indirectement – de l'activation des processus de renforcement, provoquée par la stimulation des zones érogènes des organes génitaux.
À noter par rapport à cette hypothèse que si le comportement permettant la reproduction est acquis, les caractéristiques biologiques qui permettent indirectement son apprentissage sont innées. Mais ce qui est inné serait les conditions qui rendent hautement probable l'acquisition d'un comportement permettant la reproduction, et non des motivations ou des séquences motrices neurobiologiquement préprogrammées pour le coït vaginal.
En synthèse, les processus de renforcement, les zones érogènes, ainsi que l'anatomie et la physiologie de la reproduction sont innés, et induiraient indirectement l'acquisition d'un comportement permettant la reproduction (Figure 22).
Plus précisément, les caractéristiques anatomiques et physiologiques de l'appareil reproducteur (vagin, utérus, pénis, cycles menstruels, régulation hormonale de la gamétogenèse ...) sont innées. D'autres caractéristiques innées, non liées à la reproduction, dont en particulier les processus de renforcement [1] et les zones érogènes [2], créent des conditions particulières qui induiraient l'apprentissage quasi certain de séquences comportementales de recherche et de production du plaisir physique [3]. La complémentarité des organes génitaux [4] et leur potentiel hautement érogène (orgasme) induiraient une forte probabilité d'apprentissage du coït vaginal [5].
b – Présentation détaillée du modèle comportemental
L'objectif des paragraphes ci-dessous est de présenter un modèle de l'apprentissage du comportement permettant la reproduction, qui découle de l'hypothèse principale de cette thèse. La description, chez l'Homme, de la dynamique supposée des processus et des situations, devrait ainsi permettre de bien comprendre le rôle primordial des processus de renforcement dans l'émergence et l'apprentissage du comportement permettant la reproduction.
À noter que ce travail de recherche est essentiellement destiné à vérifier l'hypothèse principale (c'est-à-dire le rôle prépondérant des processus de renforcement), et pas le modèle comportemental ci-dessous. Ce modèle est partiellement spéculatif et il existe actuellement peu de références permettant de le corroborer dans sa totalité. Il est uniquement présenté dans un but de compréhension, afin de donner un cadre global et cohérent à l'hypothèse principale. Sa vérification expérimentale sera réalisée ultérieurement.
Acquisition du comportement permettant la reproduction
En réalisant une synthèse des connaissances actuelles, on observe chez tous les mammifères une organisation sexuée innée, adaptée à la reproduction : les organes génitaux, externes comme internes, ont des structures et des fonctions spécifiques à la reproduction (pénis et vagin, testicules, ovaires, utérus…) et sont parfaitement adaptés à ce but. Au niveau physiologique, les organismes mammaliens sexués possèdent des caractéristiques qui sont également adaptées à la reproduction (sexuation, régulation hormonale de la gamétogenèse, complémentarité fonctionnelle des gamètes…). Il existe également des réflexes spécifiques (érection, lubrification, éjaculation…), précablés dans le système nerveux autonome, qui participent à la reproduction. Ces données confirment qu'il existe, au niveau anatomique et physiologique, une organisation innée, spécifique et adaptée à la reproduction (KNOBIL & NEILL 1994).
La seule composante de la reproduction qui ne soit pas innée chez tous les mammifères est, au niveau comportemental, l'acte moteur qui permet le déplacement des spermatozoïdes du méat urétral du mâle au col de l'utérus de la femelle.
En fonction des données présentées et analysées dans le chapitre précédant, ce comportement crucial, le coït vaginal, qui permet le dépôt des spermatozoïdes dans le vagin, serait chez l'Homme indirectement appris en raison des caractéristiques biologiques innées suivantes ( Figure 22, parties [1], [2] et [4], page 50 ) :
a) L'existence innée de processus de renforcement (vraisemblablement localisés dans la partie basse du faisceau médian du télencéphale : nucleus accumbens, aire tegmentale ventrale, ...).
b) L'existence innée de zones corporelles érogènes (principalement : gland du pénis et clitoris ; secondairement : lèvre, langue, seins, anus, ...).
c) L'existence innée d'organes génitaux complémentaires (pénis et vagin), ayant une haute potentialité érogène (orgasme).
Ces trois caractéristiques semblent être à l'origine de deux phénomènes particuliers : 1) principalement, l'émergence d'une dynamique comportementale particulière : le comportement érotique, et 2) secondairement, l'apprentissage d'une séquence motrice spécifique : le coït vaginal.
Émergence d'une dynamique comportementale : le comportement érotique
L'association de deux facteurs biologiques innés, les processus de renforcement [1] avec les zones corporelles érogènes [2], crée un système fonctionnel. Ce système serait à l'origine de l'émergence d'un comportement érotique : celui de la répétition des séquences motrices de stimulation des zones érogènes [3].
Cette organisation anatomique et physiologique particulière fait qu'il existerait une très forte probabilité que chaque personne, au cours de son développement et de l'interaction avec les autres, découvre les zones érogènes du corps, et, par l'intermédiaire des processus de renforcements, acquière des activités répétées de stimulation de ces zones érogènes.
Chaque enfant découvrirait progressivement (dès la première enfance (KINSEY & al. 1948), si ce n’est au cours de la vie intra-utérine (BROUSSIN & BRENOT 1995, 1996 ; MEIZNER 1987)) des stimulations corporelles et génitales qui produisent des sensations de plaisir intense. Ces premières stimulations génératrices de sensations érotiques peuvent être très diverses, et proviendraient généralement des soins parentaux, des jeux sexuels, d'une initiation par un partenaire plus expérimenté, ou de l'auto-stimulation manuelle des organes génitaux (du fait de la disposition anatomique des membres antérieurs, particulièrement propice à cette activité). Puis c'est au hasard des expériences et des apprentissages personnels (KINSEY 1948) qu'un sujet découvrirait peu à peu – ou ne découvrirait pas – les différentes pratiques et sources de plaisirs érotiques : caresses sensuelles, masturbation, baiser, activités oro-génitales, coït anal ou vaginal … Les activités érotiques seraient ainsi graduellement acquises au cours du développement, en quelques mois ou en plusieurs années, en fonction des apprentissages, de la quantité et de la qualité des expériences érotiques.
Il serait même possible de parler de comportement érotique, dans la mesure où les renforcements (perçus consciemment comme sensation de plaisir érotique) agiraient comme un principe organisateur et structurant : au cours du temps et des expériences, les activités érotiques deviendraient plus en plus typiques, élaborées, identifiées, conscientes et délibérées. À la maturité, le but recherché, les schèmes mentaux et les activités motrices seraient structurés et organisés autour d'une finalité spécifique : celle de l'obtention des sensations érotiques, dont, surtout, la jouissance orgastique.
Le plaisir érotique (ou les renforcements érotiques) pourrait ainsi être considéré comme un facteur psychique et comportemental majeur, tant dans le développement que dans la dynamique du comportement érotique.
Remarques complémentaires
Afin de compléter la description ci-dessus du comportement érotique, il est important de rappeler que les stimuli qui déclenchent le plaisir érotique sont des stimuli somatiques mécaniques qui peuvent être provoqués par des activités sans aucun lien avec la reproduction. De nombreux types de contacts corporels – qu'ils soient ou non considérés comme sexuels ou érotiques, et par différents êtres vivants ou par des objets – peuvent provoquer un plaisir physique intense.
De plus, il est très important de rappeler que ce comportement érotique, de par la dynamique des processus biologiques qui le sous-tendent, ne serait pas spécifiquement destiné à la réalisation du coït vaginal. Ce ne serait donc pas un comportement de reproduction. Il peut être qualifié d'érotique dans la mesure où la stimulation des zones érogènes provoque, en plus de l'activation des processus de renforcement, des sensations conscientes de plaisir intense, et que ce sont ces sensations psychiques érotiques qui semblent être le but conscient délibérément recherché, et non la reproduction.
Enfin, il est également très important de noter que ce comportement érotique, de par la dynamique des processus biologiques qui le sous-tendent, n'existerait pas à la naissance, ni au niveau moteur, ni au niveau psychique. Il n'existerait pas à la naissance ni de besoin, de désir ou de pulsion érotique, ni d'activité érotique spontanée (WUNSCH & BRENOT 2004 ; 2005). Ces activités érotiques ne seraient pas instinctuelles, c'est-à-dire produites par des structures neurales motrices précablées en réponse à des stimuli-signaux innés et spécifiques de l'érotisme (ou de la reproduction ou de la "sexualité"). Ce ne serait qu'à partir du moment où une personne a ressenti du plaisir érotique et à compris par quelle action il a été provoqué, qu'il pourrait exister un désir érotique et une motivation à reproduire ces actions si intensément hédoniques. Ce qui semble être déterminant dans le développement du comportement érotique, ce ne serait pas tant le genre du partenaire, l'âge ou le type de l'activité érotique, mais plutôt la découverte de la première sensation érotique, la compréhension de la cause de ce plaisir, puis la répétition et la recherche de nouvelles sensations de ce plaisir si intense.
Ainsi, en fonction du contexte socioculturel et des expériences singulières de chaque personne, le développement du comportement érotique pourrait commencer dès l'enfance, à l'adolescence ou à l'âge adulte, par des activités autosexuelles, hétérosexuelles, homosexuelles ou bisexuelles. Les premières activités peuvent être n'importe lesquelles, caresses ou stimulations manuelles, masturbation, baiser, activités oro-génitales ou coït, en fonction de la situation et de l'expérience d'un éventuel partenaire initiateur. L'influence des facteurs culturels serait très importante : par exemple, en Occident, l'homophobie et la valorisation sociale du couple hétérosexuel, induisent un contexte où les activités sexuelles entre un homme et une femme seront majoritaires. Dans ce contexte, les renforcements érotiques ont lieu essentiellement entre personnes de sexe opposé, ce qui induirait ainsi le développement d'une sexualité majoritairement hétérosexuelle. Quelle que soit la société, l'enfant, l'adolescent, puis le jeune adulte observe, imite, apprend et se conforme aux mœurs sexuelles en usage dans son groupe social. Ainsi, quel que soit le contexte socioculturel, la grande majorité des renforcements érotiques ne concernent que les activités de stimulation des zones érogènes socialement acceptées, ce qui entraînerait leur répétition et ainsi la conformité du comportement sexuel aux normes sociales.
Apprentissage d'une séquence motrice spécifique : le coït vaginal
Par rapport à l'apprentissage de la séquence du coït vaginal, il conviendrait peut être de distinguer deux contextes : un contexte initial d'ignorance du coït vaginal, correspondant à l'époque préhistorique des tous premiers Hommes, et un contexte postérieur, correspondant à la connaissance de ce coït.
Dans le premier contexte, comme on ignore ses caractéristiques exactes, nous allons nous mettre dans la situation la plus défavorable et supposer que la séquence du coït vaginal n'était vraisemblablement pas connue. Même dans ce cas, il semble très probable que le comportement de reproduction puisse être acquis. La découverte et l'apprentissage du coït vaginal ont pu se faire de la manière décrite dans les paragraphes ci-dessous.
Dans le second contexte, qui représente la quasi-totalité de l'histoire humaine, le coït vaginal est déjà connu. Il serait appris soit par l'observation des activités coïtales d'autres personnes, soit par une initiation par un partenaire plus expérimenté, soit par la transmission théorique de cette connaissance par un moyen oral, écrit ou pictural. Dans ce deuxième contexte, les caractéristiques biologiques spécifiques à l'origine de l'acquisition du coït vaginal (principalement les processus de renforcement associés aux zones érogènes génitales) ne joueraient plus aucun rôle dans la découverte de cette séquence motrice, mais resteraient essentielles pour sa répétition.
Comment, à l'époque des tous premiers Hommes, en l'absence d'instinct, de connaissances et d'informations, le coït vaginal peut-il être acquis ? D'après le modèle proposé (voir Figure 22), la découverte et l'apprentissage du coït vaginal semblent possible et hautement probable, et ont vraisemblablement dus se réaliser à l'époque originelle de la manière suivante :
Durant le comportement érotique, au cours de la recherche de plaisirs physiques, l'existence d'organes génitaux complémentaires (pénis et vagin) ayant des zones hautement érogènes [4] serait une particularité qui induirait une forte probabilité que le coït vaginal devienne une des pratiques érotiques préférentielles [5]. ( Figure 22, parties [4] et [5], page 50 )
Le fait que la stimulation des organes génitaux est généralement susceptible de provoquer un plaisir érotique très intense, dont l'orgasme, serait un des principaux facteurs facilitant l'acquisition et la répétition du coït vaginal. Cette caractéristique ferait que les activités érotiques sont principalement centrées sur les organes génitaux (masturbation réciproque, fellation, cunnilingus, etc.), augmentant ainsi la probabilité de découverte du coït vaginal. De plus, la curiosité, la recherche de la nouveauté et les capacités cognitives seraient également des facteurs augmentant la probabilité de découverte de cette séquence. Ainsi, entre la maturation des capacités motrices, vers 3-4 ans, et la puberté, il existe une période d'une dizaine d'années d'expérimentations érotiques, vraisemblablement plus que suffisante pour découvrir la séquence du coït vaginal. Cette séquence serait ainsi acquise au cours du développement et de la diversification des activités répétées de stimulations des zones érogènes.
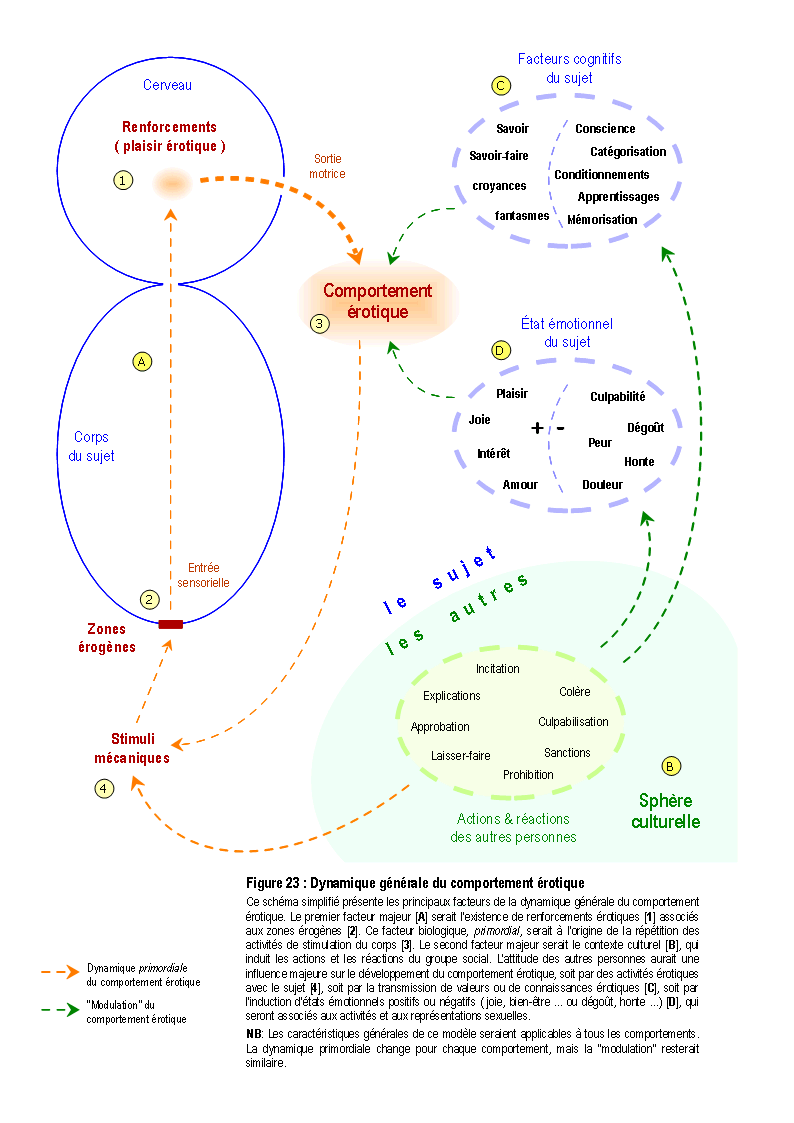
Par ailleurs, pour l'homme, la position anatomique du gland et du prépuce à l'extrémité du pénis ainsi que leur physiologie sensorielle font que la pénétration, vaginale mais également anale ou buccale, serait une des activités qui provoque le plaisir érotique le plus intense. Pour la femme, la sensibilité propre du vagin et sa relation étroite avec le clitoris, font que la pénétration vaginale serait également une des activités qui provoquerait le plaisir érotique le plus intense. De plus, la lubrification vaginale, la possibilité d'un important contact corporel (dans certaines positions) ainsi que la possibilité d'autres activités érotiques simultanées (caresses sensuelles, baiser ...), feraient que le coït vaginal deviendrait une des activités érotiques préférées, favorisant ainsi la fécondation et la reproduction nécessaire à la survie de l'espèce.
En conclusion, il semblerait donc que le coït vaginal, nécessaire à la reproduction et à la survie de l'espèce humaine, dépende d'un apprentissage spécifique, généralement mais pas obligatoirement effectué au cours de l'apprentissage des différentes activités érotiques. Dans l'espèce humaine et peut-être chez d'autres primates anthropoïdes, la reproduction semblerait être un heureux aléa, conséquence presque fortuite de la recherche du plaisir physique.
Figure 23
Autres facteurs intervenants dans le comportement de reproduction
À cette dynamique comportementale de base, où les processus de renforcement seraient le facteur primordial, interviendraient, en plus, d'autres facteurs biologiques et culturels ( Figure 23 ).
Ces facteurs n'auraient pas tous la même importance dans la sexualité. Les facteurs les plus primaires et les plus importants seraient les processus de renforcement ( Figure 23, partie [1], et Figure 22, partie [1] ), ainsi que les hormones et les phéromones sexuelles. Les facteurs secondaires seraient les processus émotionnels et cognitifs ( Figure 23, partie C & D ). Enfin, les facteurs tertiaires seraient les facteurs sociaux et culturels ( Figure 23, partie B ).
Facteurs primaires : Hormones & phéromones sexuelles
L'hypothèse de l'importance primordiale des processus de renforcement n'est absolument pas incompatible avec un rôle comportemental des hormones et des phéromones sexuelles. Certains effets hormonaux et phéromonaux mis en évidence par l'expérimentation animale existent très certainement chez l'Homme. Néanmoins ces effets seraient, comparés aux effets majeurs des processus de renforcements érotiques, relativement plus faibles.
Vraisemblablement, les processus hormonaux et phéromonaux auraient une action modulatrice. Par exemple les hormones sexuelles (principalement la testostérone) augmenteraient à la puberté l'intensité du désir et du comportement érotique, tandis que les phéromones sexuelles pourrait augmenter la proportion d'activités hétérosexuelles, favorisant ainsi la reproduction.
Remarque : à noter qu'il semble très important de bien distinguer les effets anatomiques, physiologiques et comportementaux les uns des autres. Par exemple, bien que les hormones sexuelles ont, chez l'Homme, un effet majeur au niveau anatomique et physiologique de l'appareil reproducteur, cela n'impliquerait absolument pas qu'elles doivent également avoir un effet majeur sur le comportement qui permet la reproduction.
Facteurs secondaires : Émotion & Cognition
De plus, les processus émotionnels (plaisir, joie, peur, dégoût, affects, attachement ... Figure 23, partie D ) et cognitifs (conditionnement pavlovien et opérant, habituation, extinction, associations, apprentissages, mémorisation, planification, catégorisation ... Figure 23, partie C ) complexifieraient tant les séquences comportementales que les états affectifs, et, surtout, permettraient une dimension psychique et consciente aux activités érotiques.
Enfin, les processus cognitifs les plus complexes seraient à l'origine d'une dimension culturelle de la sexualité, en permettant l'élaboration d'éléments abstraits (représentations, valeurs, croyances ...) qui transformeraient la simple étreinte des corps en actes éminemment chargés de sens et de symboles.
Remarque : à noter que la majorité de ces facteurs émotionnels et cognitifs ne seraient pas indispensables à la réalisation du comportement permettant la reproduction. Comme ils existent dans l'organisme humain, ils agissent forcément sur le développement et la dynamique de tous les comportements et états psychiques, dont le comportement et les affects sexuels. Mais, en théorie, la plupart pourraient être supprimés sans affecter la reproduction de l'organisme.
Facteurs tertiaires : facteurs sociaux & culturels
Les facteurs sociaux et culturels ( Figure 23, partie B ), qui se traduisent concrètement par des attitudes et des actions particulières des personnes du groupe social, seraient les facteurs les moins importants. Ils n'agiraient que sur ce qui est biologiquement possible. Le contexte social et culturel peut interdire ou valoriser telles ou telles pratiques, valeurs ou croyances sexuelles, il peut induire le dégoût des organes génitaux, la crainte du péché de luxure ou l'élaboration du concept de "sexualité", mais il ne pourrait modifier le fonctionnement biologique des processus neurobiologiques primordiaux impliqués dans la reproduction et les renforcements érotiques.
Pathologie : Addictions sexuelles
Par ailleurs, ce modèle permettrait d'expliquer l'étiologie des principaux troubles de la sexualité (WUNSCH & BRENOT 2005), dont par exemple l'étiologie de l'addiction sexuelle.
Les processus neurobiologiques impliqués dans les renforcements érotiques inconscients et/ou les perceptions érotiques conscientes seraient – au moins partiellement – les mêmes que ceux impliqués dans la dépendance aux drogues opioïdes.
Il est possible qu'au-delà d'un certain seuil d'activité et d'intensité, l'organisme deviendrait dépendant aux sensations érotiques, éventuellement par l'intermédiaire des opioïdes endogènes. Ce phénomène neurobiologique pourrait être une des principales causes à l'origine des addictions sexuelles (REYNAUD 2005 ; AGMO & BERENFELD 1990).
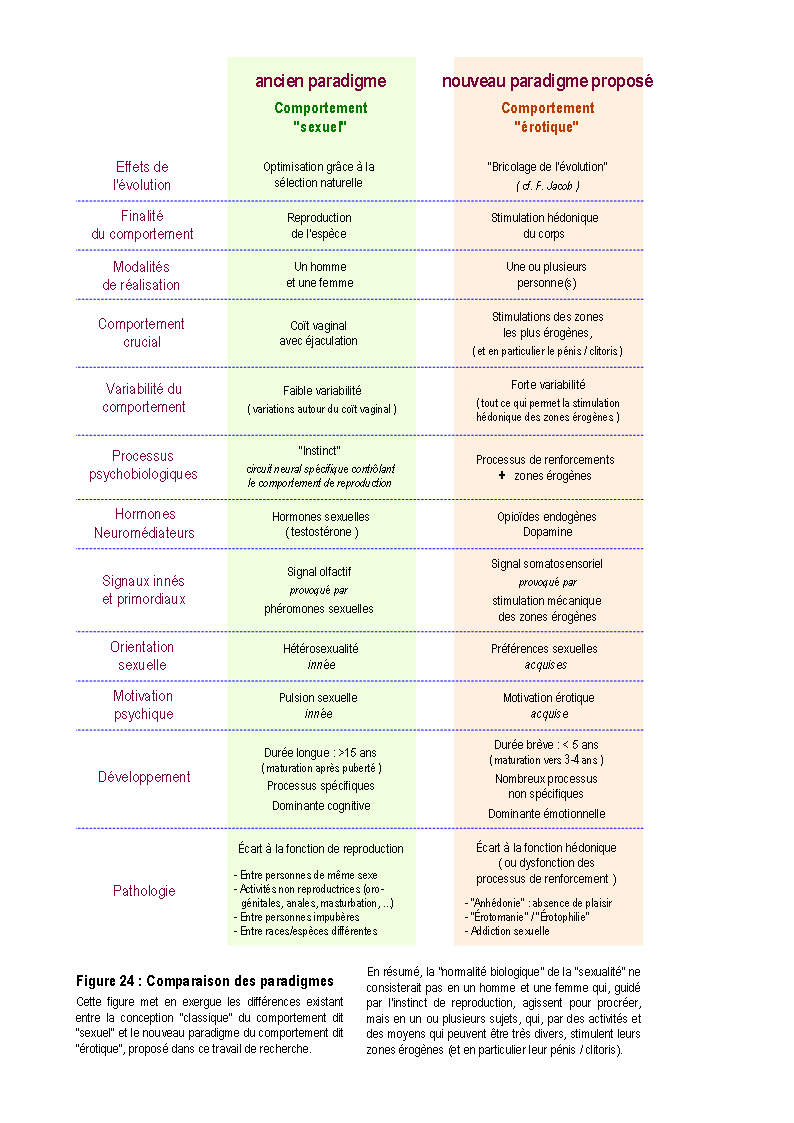
c – Comparaison entre les modèles de la reproduction
Pour terminer la présentation de ce modèle – qui correspond à un nouveau paradigme (KUHN 1983) de compréhension et d'explication du comportement de reproduction – est donné un tableau récapitulatif, qui, en opposant un à un tous les éléments clés, met en exergue les importantes différences qui existent entre ce modèle et le paradigme classique, innéiste, du comportement de reproduction humain (Figure 24).
Le modèle classique explique la sexualité humaine principalement à partir d'un "instinct" de la reproduction tandis que ce modèle est basé sur la prépondérance fonctionnelle des processus de renforcement érotique. Depuis les années 1960 / 70, on observe une évolution des théories sexologiques de l'ancien vers le nouveau modèle, en particulier par la "normalisation" de nombreuses pratiques qui étaient autrefois considérées comme pathologiques. Comme principaux exemples, on peut donner : la quête de plaisirs érotiques diversifiés, la masturbation, le coït anal, les activités oro-génitales et l' "homosexualité".
À noter que ce modèle, chez l'Homme, de l'apprentissage du comportement permettant la reproduction, est en parfaite cohérence avec la théorie de l'évolution de François Jacob (prix Nobel de médecine en 1965). Sa théorie est généralement désignée par la métaphore du « bricolage de l'évolution » (JACOB 1977, JACOB 1981). Cette théorie, basée sur l'observation des bizarreries et des imperfections de structure et de fonction du monde vivant, suppose que l'effet majeur de la sélection naturelle n'est pas l'optimisation [4] mais plutôt l'élimination des organismes non viables. C'est-à-dire passer de l'idée que tout ce qui n'est pas optimisé est évolutivement impossible à l'idée que tout ce qui survit – peut importe la manière – est possible.
d – Pansexualité potentielle
Enfin, s'il faut désigner et nommer, chez l'Homme, ce modèle de l'apprentissage du comportement permettant la reproduction, l'expression la plus appropriée serait celle de la "pansexualité potentielle ".
En effet, on observe que l'association entre les caractéristiques neurobiologiques objectives du comportement érotique (processus de renforcement, zones érogènes, orgasme ...) avec les caractéristiques subjectives des processus cognitifs (représentations, croyances, symbolisation ...), rend possible le fait que potentiellement tout peut devenir "sexuel".
Le caractère potentiel de la sexualité humaine serait double :
– D'une part, l'activité érotique objective serait potentielle : l'activité érotique en général, et chaque type d'activités érotiques en particulier, ne se développeraient et ne se diversifieraient que si le milieu environnant et/ou le contexte culturel le permet.
– D'autre part, la représentation subjective et culturelle de la "sexualité" serait elle aussi potentielle : dans un premier temps, le concept de "sexualité" n'existerait que s'il est conceptualisé comme tel ; et, ensuite, ne deviendrait "sexuel" que ce qui est nommément désigné et spécifié comme "sexuel".
Le caractère pan-sexuel (du grec pan, pantos signifiant "tout") de la sexualité humaine serait également double :
– D'une part, l'activité érotique objective serait pan-sexuelle : quasiment tout être vivant (et tout particulièrement les mammifères familiers de taille et/ou d'apparence humaine) voire des éléments matériels ou abstraits pourraient être l'objet d'activités érotiques diverses et/ou pourraient être source d'affects érotiques.
– D'autre part, la notion subjective et culturelle de "sexualité" serait également pan-sexuelle : l'être humain pourrait attribuer la caractéristique de "sexualité" à quasiment tout élément matériel ou abstrait.
À noter que cette nature pansexuelle potentielle de la sexualité ne serait pas absolue. Les études de Pavlov et de Skinner, entre autres, ont montré que tous les types de renforcements, d'associations ou de conditionnements ne sont pas réalisables. Néanmoins, ce qui reste biologiquement possible est considérable, comme en témoigne toute la diversité des sexualités observées dans les centaines de sociétés humaines connues (FORD & BEACH 1965).
À noter également que chaque personne n'est en général pas consciente de sa potentialité pansexuelle. Il semble que subjectivement chaque sujet ne puisse considérer comme "sexuel" que ce qui est aux "normes" de sa culture, de ses connaissances, et, surtout, de son expérience concrète et singulière.
Si cette proposition de la "pansexualité potentielle ", explicative du comportement permettant la reproduction, est juste, elle permettrait d'expliquer la diversité des sexualités humaines observées à travers les âges et les cultures.
e – Possible extension du modèle et perspectives de recherche
Le modèle présenté dans les sections précédentes semble pouvoir être généralisé à d'autres comportements fondamentaux pour la survie de l'individu et de l'espèce.
En effet, il apparaît que la plupart des zones érogènes correspondent à une structure particulière, à savoir du tissu muco-cutané, qui est un tissu de transition entre la peau externe et les muqueuses internes. Cette peau particulière est caractérisée par une moindre épaisseur du derme et les structures neurales sensitives sont plus proches de l'épiderme que dans les autres types de peau (glabre ou poilue). Les zones érogènes constituées de tissu muco-cutané sont le pénis/clitoris, le prépuce, la partie externe de la vulve, la peau périanale, les mamelons et les lèvres (WINKELMANN 1959). On remarque que quasiment tous les orifices de l'organisme qui sont impliqués dans les entrées et les sorties de matières possèdent un tissu muco-cutané, vraisemblablement en relation avec les processus de renforcement. Ce système, constitué par le tissu muco-cutané associé aux processus de renforcement, semblerait avoir pour fonction de renforcer tous les comportements d'excrétion ou d'incorporation d'éléments matériels (fèces, lait, eau, aliments, gamètes ...) qui sont essentiels au fonctionnement de l'organisme.
Par exemple, l'allaitement, qui est une séquence cruciale du comportement maternel, dépendrait d'une dynamique similaire à celle qui vient d'être présentée pour le comportement érotique. En effet, des données expérimentales montrent que chez la rate la stimulation des mamelons provoque un renforcement majeur, supérieur à celui induit par la cocaïne (FERRIS & al. 2005). De plus, si l'on anesthésie le museau des ratons (STERN & JOHNSON 1990) ou les mamelles de la rate (STERN & al. 1992), cette dernière cesse l'allaitement. Ces données, similaires à celles obtenues chez d'autres femelles de mammifères (cochon d'Inde, lapin, porc, brebis, poney et baleine), suggèrent que c'est le renforcement provoqué par la stimulation des mamelons qui est le principal facteur à l'origine de la répétition de l'activité d'allaitement. Par ailleurs, certaines études ethnologiques apportent des données complémentaires intéressantes : on observe dans les tribus de Nouvelle-Guinée, qui sont considérées comme ayant un mode de vie proche des premiers Hommes, que les mères ressentent un orgasme lors de l'allaitement et qu'elles donnent le sein afin de ressentir ces sensations hédoniques (HERDT 1990 ; voir également la discussion relative aux zones érogènes, page 121). Plus intéressant encore, et de manière similaire aux autres primates, on observe que les mères après le sevrage n'alimentent plus de manière spontanée leur progéniture (MARSHALL 1981), ce qui suggère que le renforcement provoqué par la tétée serait le seul facteur "instinctif" à l'origine d'un comportement de nourrissage. Toutes ces données suggèrent que chez les mammifères le renforcement provoqué par la stimulation du tissu muco-cutané du mamelon serait le facteur primordial, essentiel à l'apparition et à la répétition du comportement d'allaitement. Après le sevrage, et donc en l'absence du renforcement provoqué par la stimulation des mamelons, il n'existerait plus de facteurs, autres que culturels chez l'Homme, à continuer l'alimentation de la progéniture.
En conclusion, il semble exister un système somatosensoriel particulier et fondamental, à l'interface entre l'intérieur et l'extérieur de l'organisme, qui participerait à l'émergence et à l'acquisition des comportements impliqués dans le contrôle des flux d'énergie et de matières (gamètes, nourriture, déchets), nécessaires au fonctionnement de l'organisme. Ces comportements cruciaux (excrétion, alimentation, allaitement, coït ...) sont absolument nécessaires à la survie de l'individu et de l'espèce.
À noter que dans ce cadre, le plaisir érotique et l'orgasme ne pourraient plus être qualifiés de "sexuels", dans la mesure où leur fonction et leur dynamique ne seraient plus en relation exclusive avec le comportement sexuel et les régions sexuées de l'organisme.
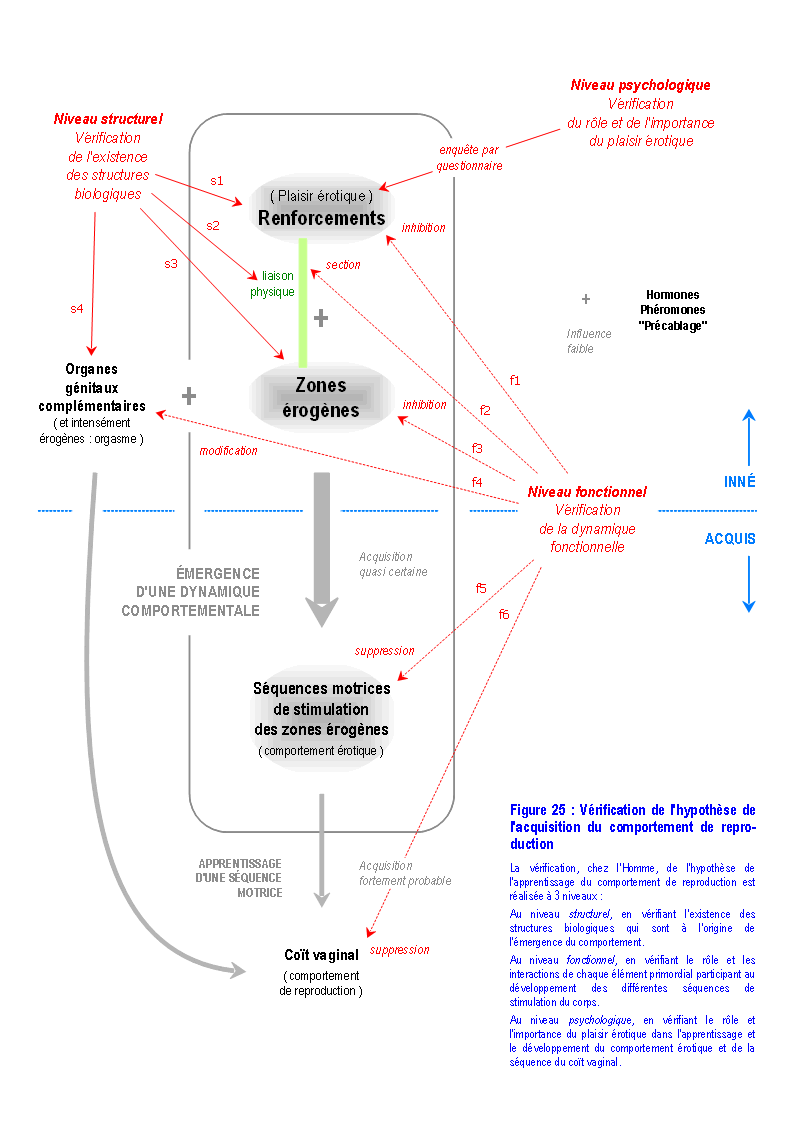
Si des études complémentaires confirment l'existence de ce système fonctionnel (i.e. tissus muco-cutané associé aux processus de renforcement), la présence des zones érogènes primaires au niveau des principaux orifices de l'organisme aurait ainsi une signification fonctionnelle adaptative. Quant au comportement érotique, sa dynamique fonctionnelle serait précisée : ce comportement apparaîtrait, se développerait et s'organiserait en fonction de la recherche de stimulations optimales du tissus muco-cutané. La mise en évidence de cette dynamique permettrait de comprendre la raison pour laquelle les activités érotiques se développent essentiellement autour des principaux orifices de l'organisme, n'ont pas de lien direct avec la reproduction et semblent a priori tellement "contre-nature".
Quant à l'ensemble des comportements humains (maternel, agression, alimentation ...), leur développement et leur dynamique générale seraient similaires à celles du comportement érotique (cf. le résumé synoptique dans la Figure 23, page 55 ). Il existerait pour chaque comportement quelques processus primordiaux qui seraient à l'origine d'une dynamique particulière. Ces processus seraient différents pour chaque comportement. Puis, la dynamique primordiale de chaque comportement serait "modulée" par les processus émotionnels et cognitifs, ainsi que par les actions et les réactions des autres personnes du groupe social. Chaque comportement final adulte résulterait d'une interaction de nombreux facteurs, mais les processus neurobiologiques primordiaux seraient les seuls facteurs cruciaux et déterminants, à l'origine du développement et de la spécificité de chacun de ces comportements. Le principal enjeu de la recherche comportementale serait l'identification et la caractérisation de ces processus neurobiologiques primordiaux.
f – Méthode de vérification de l'hypothèse principale
La vérification complète et exhaustive d'un nouveau modèle comportemental nécessite de nombreuses vérifications systématiques de tous les aspects cruciaux de ce modèle.
Par rapport au modèle de l'apprentissage du comportement permettant la reproduction (cf. Figure 22), sa vérification systématique nécessite de réaliser plusieurs expérimentations à trois niveaux différents : au niveau structurel, au niveau fonctionnel, et au niveau psychique (Figure 25).
Figure 25
Niveau structurel
Au niveau structurel, l'objectif des premières étapes de la vérification est de réaliser des expérimentations qui permettent de valider l'existence des différentes structures constituant le système neural à l'origine du comportement permettant la reproduction ( Figure 25, partie [s1], [s2], [s3] et [s4] ).
C'est-à-dire réaliser des expérimentations qui vérifient : l'existence de processus de renforcement spécifiquement activés lors des stimulations des zones érogènes [s1] ; vérifier l'existence et la nature des liaisons fonctionnelles entre les zones érogènes et les processus de renforcement [s2] ; vérifier l'existence, le nombre, la localisation, le développement et les caractéristiques des différentes zones érogènes [s3] ; et enfin vérifier que le pénis, le clitoris et/ou le vagin sont bien les organes à l'origine du renforcement le plus intense, c'est-à-dire vraisemblablement qu'ils seraient les structures clés permettant de provoquer l'orgasme [s4].
Niveau fonctionnel
Au niveau fonctionnel, l'objectif des principales étapes de la vérification est de réaliser des expérimentations qui permettent de valider les phases théoriques du développement et la dynamique supposée du comportement permettant la reproduction ( Figure 25, partie [f1], [f2], [f3], [f4], [f5] et [f6] ).
C'est-à-dire réaliser des expérimentations qui vérifient, entres autres, que l'inhibition sélective dans le fœtus des processus de renforcement fonctionnellement liés aux zones érogènes devrait supprimer le développement du comportement érotique [f1] ; que la section dans le fœtus de la liaison entre une zone érogène et les processus de renforcement devrait supprimer le développement d'un comportement de stimulation de cette zone [f2] ; que l'inhibition sensorielle transitoire d'une zone érogène devrait supprimer le développement d'un comportement de stimulation de cette zone jusqu'à la période de suppression de cette inhibition [f3] ; que la modification de l'intensité du renforcement procuré par les organes génitaux, par exemple la minimalisation du renforcement, devrait provoquer un relatif désintérêt de la zone érogène génitale [f4] ; que la suppression des stimulations érotiques par des partenaires devrait supprimer le développement d'un comportement érotique orienté vers les autres [f5] ; et enfin, que la suppression juste du coït vaginal par les partenaires devrait induire le développement d'un comportement érotique où le sujet aurait de nombreux désirs d'activités érotiques, mais aucun relatif au coït vaginal [f6]
Niveau psychique
Au niveau psychique, l'objectif des dernières étapes de la vérification est de réaliser des expérimentations qui permettent de valider l'existence de relations entre les processus neurobiologiques supposés et les perceptions qu'ils devraient engendrer au niveau psychique ( Figure 25, partie [Niveau psychologique] ).
C'est-à-dire réaliser des expérimentations qui vérifient, entres autres, qu'en l'absence de toutes activités ou états érotiques il ne devrait exister aucun désir érotique ; que le désir psychique pour une activité érotique précise ne devrait exister qu'à partir de la période où cette activité à été vécue de manière positive pour la première fois ; et que le plaisir érotique devrait être perçu comme étant le principal facteur de l'économie psychique sexuelle.
Problèmes théoriques, pragmatiques et éthiques
Il est donc nécessaire de réaliser un grand nombre d'expérimentations, qui sont pour certaines longues et/ou complexes, afin de vérifier de manière systématique le nouveau modèle de l'apprentissage du comportement permettant la reproduction.
De plus, il existe également des limites dues au manque actuel de connaissances concernant le fonctionnement détaillé des processus de renforcement et les caractéristiques précises des zones érogènes. Ces lacunes nécessiteraient de réaliser plusieurs expérimentations préliminaires, afin par exemple de préciser en détail les mécanismes à l'œuvre, les neuromédiateurs, l'architecture et la localisation exacte des diverses structures impliquées dans les renforcements érotiques.
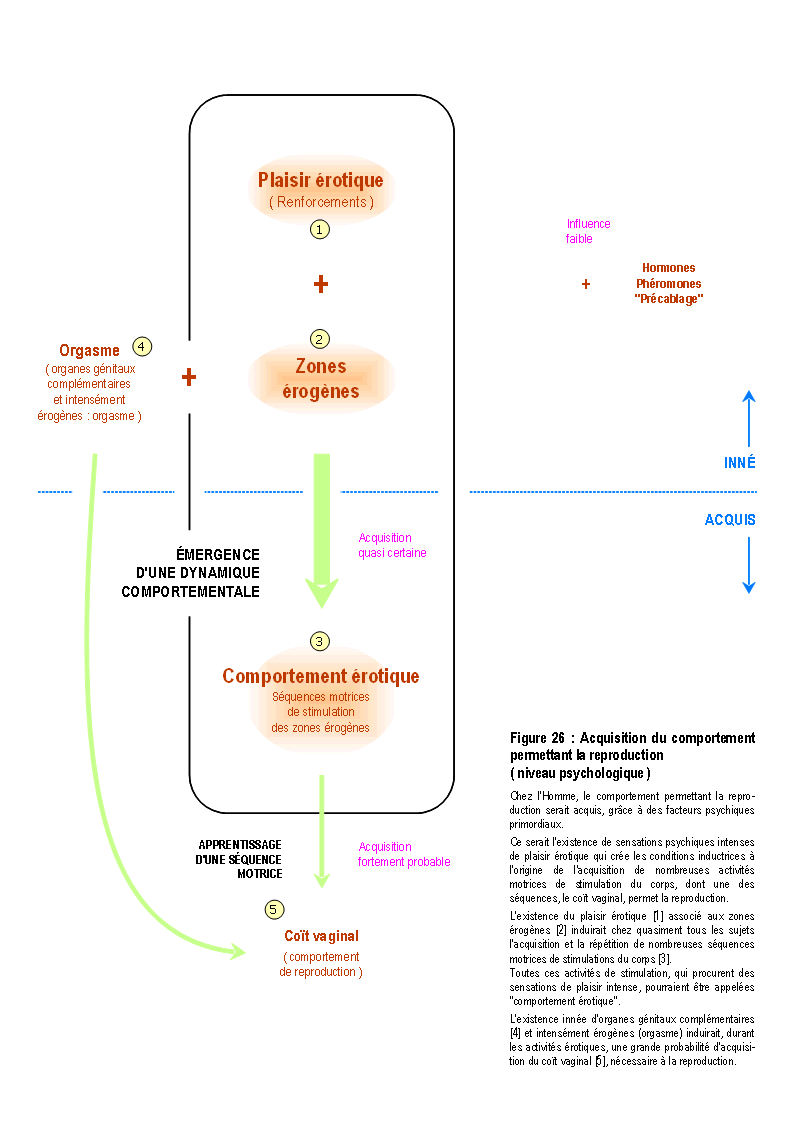
Enfin – et surtout – il est impossible pour des raisons éthiques de réaliser chez l'Homme la plupart de ces expérimentations. Tout ces problèmes rendent difficile la vérification du modèle.
Cependant, une possibilité alternative a été explorée, consistant en l'utilisation des situations "naturelles" particulières, proches des conditions expérimentales exposées dans les paragraphes ci-dessus. Par exemple l'anhédonie (BOURGEOIS 1999), certaines atteintes médullaires pathologiques ou traumatiques (SOULIER 2001) ou des contextes culturels défavorables à l'expression de certaines activités sexuelles (FORD & BEACH 1965). Néanmoins la plupart de ces situations ne permettent pas de recueillir des données fiables, en raison de l'existence de nombreux paramètres non contrôlés, d'atteintes pathologiques difficiles à évaluer, de vraisemblables réorganisations cérébrales ou d'influences culturelles impossibles à mesurer.
Pour toutes ces raisons, il a été impossible de réaliser la plupart des expérimentations nécessaires. La vérification de l'hypothèse principale a donc été finalement effectuée au niveau psychologique, au moyen d'un questionnaire.
L'hypothèse principale étant une hypothèse neurobiologique, il a été nécessaire, afin de déterminer les caractéristiques psychiques à investiguer au moyen du questionnaire, de la transposer au niveau psychologique.
g – Transposition de l'hypothèse du niveau neurobiologique au niveau psychologique
L'hypothèse de l'acquisition du comportement permettant la reproduction peut être décrite à deux niveaux d'organisation : le niveau biologique (ou neurobiologique) et le niveau psychologique.
Au niveau neurobiologique, le facteur clé de l'apprentissage du comportement érotique seraient les processus de renforcement érotique. Au niveau psychologique, il semblerait que cette activité des renforcements érotiques soit perçue comme une sensation consciente de plaisir érotique.
L'hypothèse principale H, sous réserve qu'il existe une relation étroite entre renforcement érotique et plaisir érotique conscient, peut être exprimée aux deux niveaux d'organisation :
Figure 26
H : Niveau (neuro)biologique : L'acquisition du comportement permettant la reproduction dépend – principalement mais indirectement – de l'activation des processus de renforcement, provoquée par la stimulation des zones érogènes des organes génitaux (Figure 22, page 50).
Hy : Niveau psychologique : L'acquisition du comportement permettant la reproduction dépend – principalement mais indirectement – de la recherche de sensations conscientes de plaisir érotique, provoquées par la stimulation des zones érogènes des organes génitaux. (Figure 26)
L'objectif de la vérification expérimentale est d'examiner, au niveau psychologique, le rôle et l'importance de la sensation de plaisir érotique dans la dynamique et l'apprentissage du comportement de reproduction.
La vérification de l'hypothèse Hy est réalisée à partir de 3 hypothèses intermédiaires clés. Ces 3 hypothèses ont été élaborées de manière à pouvoir réfuter l'hypothèse Hy , c'est-à-dire en recherchant les conditions expérimentales qui sont les plus susceptibles de montrer que l'hypothèse est fausse.
C'est-à-dire, si le plaisir érotique est le facteur majeur à l'origine des activités érotiques (hypothèse Hy ), on devrait absolument trouver que :
– Hypothèse intermédiaire Hy1 : Le plaisir érotique doit être un des plaisirs les plus intenses que l'organisme peut éprouver.
Dans le cas contraire, afin de ressentir des plaisirs intenses, les personnes préféreront certainement réaliser d'autres activités que les activités érotiques, ce qui compromettrait la reproduction et donc la survie de l'espèce.
– Hypothèse intermédiaire Hy2 : Les zones érogènes des organes génitaux (pénis/clitoris, vagin) doivent être à l'origine d'un des plaisirs érotiques les plus intense.
Dans le cas contraire, les personnes préféreront par exemple juste s'embrasser, ce qui compromettrait la reproduction.
– Hypothèse intermédiaire Hy3 : Le plaisir érotique provoqué par un partenaire doit être supérieur à celui provoqué par autostimulation.
Dans le cas contraire, les personnes préféreront uniquement se masturber, ce qui, là encore, compromettrait la reproduction.
Si ces 3 conditions sont vérifiées, le plaisir érotique (et donc indirectement les processus de renforcement) serait un des facteurs psychiques majeurs à l'origine du développement et de la dynamique du comportement érotique, ainsi que de l'acquisition du coït vaginal reproducteur.
Le questionnaire permettant la vérification expérimentale de l'hypothèse a donc été élaboré de manière à recueillir principalement des perceptions psychiques relatives aux activités et aux sensations érotiques, dans l'objectif de vérifier ces 3 hypothèses intermédiaires.
 |
3 – Matériel & Méthode
a – Matériel
Le questionnaire utilisé dans cette étude a été spécialement créé pour répondre aux spécificités de cette recherche. La première version du questionnaire, conçue pour une recherche exploratoire, contenait près d'une centaine de questions couvrant tous les aspects de l'hypothèse principale. Les réponses d'une trentaine de personnes, contactées dans l'entourage des auteurs, ont permis d'élaborer la version quasi finale du questionnaire à partir de l'analyse du contenu, de l'analyse des problèmes rencontrés, des commentaires des participants et des résultats obtenus.
L'objectif du questionnaire est d'évaluer le rôle et l'importance, au niveau psychique, du plaisir érotique dans la dynamique du comportement sexuel. Pour atteindre cet objectif, le questionnaire contient plusieurs groupes de questions : A] un premier groupe concerne l'évaluation des sensations érotiques, par des questions principales, relatives à la prépondérance du plaisir érotique dans les activités sexuelles (questions 13, 16 & 18, associées aux questions 1, 17 & 19), et des questions secondaires donnant des compléments d'informations sur le plaisir érotique (questions 4, 12, 14 & 15) ; B] un second groupe de questions est destiné à recueillir des informations qui permettent d'affiner les analyses des réponses relatives à l'importance du plaisir érotique : une question concernant les facteurs à l'origine des activités sexuelles (question 10), des questions relatives au profil des activités sexuelles (questions 2, 3, 5, 6, 7, 8 & 9), ainsi que des questions donnant des informations complémentaires diverses (questions 11, 20, 21 & 22) ; et C] un troisième groupe de questions est destiné à recueillir des informations permettant d'exclure de l'analyse principale les sujets ayant des troubles perturbant leurs activités sexuelles (questions 23, 24, 25 & 26).
En général, les questions comportent plusieurs items, tels par exemple ceux donnés ci-dessous, et il était demandé aux participants d'indiquer par des notes (de 0 à 9) l'intensité du plaisir ressentit pour chaque item.
…… Le plaisir gustatif (aliments, boissons, friandises, etc.) le plus intense que j'ai vécu
…… Le plaisir olfactif (odeurs, parfums, senteurs, etc.) le plus intense que j'ai vécu
…… Le plaisir auditif (sons, musique, chants, etc.) le plus intense que j'ai vécu
etc.
La finalisation et la vérification de la faisabilité de la version quasi finale du questionnaire ont été réalisés par l'intermédiaire du réseau Internet. Trois versions successives du questionnaire ont été testée et améliorées en fonction des réponses et des commentaires des internautes, ainsi que des analyses des résultats intermédiaires. La version finale du questionnaire est présentée en annexe, à la page 178.
Analyses statistiques. Les analyses statistiques, les tableaux et les figures ont été réalisée avec SPSS 13.0 et SigmaStat 3.11 pour Windows. Sauf indication contraire, le seuil de significativité à été fixé à 0,05.
b – Procédure
Le questionnaire a été mis en ligne à l'adresse http://sexualite.ouvaton.org/questionnaire.htm [5]. Des messages invitant les internautes à y répondre on été postés sur des forums dédiés à l'information sexuelle (par exemple www.doctissimo.fr) et sur le site de l'Observatoire International du Couple (www.couple.asso.fr). Le questionnaire est resté en ligne durant toute l'année universitaire 2005 / 2006.
Par ailleurs, une autre passation à été réalisée en décembre 2005 auprès d'une quinzaine d'étudiants en médecine à Bordeaux. Les étudiants étaient dans une grande salle, suffisamment loin les uns des autres pour créer un climat d'intimité et d'anonymat.
Tous les participants ont été informés que l'enquête concernait le comportement sexuel, qu'elle était totalement anonyme et basée sur le volontariat, d'une durée d'environ 30 minutes et qu'ils pouvaient arrêter la passation en cours s'ils ne désiraient plus participer à l'enquête.
c – Participants
Les 749 personnes francophones (398 femmes, 349 hommes et 2 transsexuels) qui ont participé à l'enquête par questionnaire se répartissent de la manière suivante :
Nombre de participants : 749 100 %
Cas inclus dans l'étude : 430 57 %
Femmes : 223 30 %
Hommes : 205 27 %
Cas analysés à part : 242 32 %
Cas inutilisables : 79 10 %
79 personnes ont rendu des questionnaires inexploitables à causes d'erreurs diverses. 240 personnes ont déclaré différents problèmes qui perturbent actuellement leur sexualité et ont donc été analysées à part. Les 431 personnes restantes, incluses dans les analyses principales, ont été répartit dans 20 différents groupes, en fonction principalement du sexe, de l'orientation de leurs activités sexuelles et de certaines caractéristiques liées à ces activités (absence de masturbation, d'orgasme ou d'activités sexuelles avec un partenaire, etc.).
Finalement, seul 5 groupes contiennent suffisamment de cas pour être analysés statistiquement :
Femmes hétérosexuelles ( F_Hété ) : 135 cas soit 18 % des 749 questionnaires recueillis
Hommes hétérosexuels ( H_Hété ) : 114 cas 15 %
Femmes bisexuelles ( F_Bi ) : 58 cas 7,7 %
Hommes bisexuels ( H_Bi ) : 64 cas 8,5 %
Hommes homosexuels ( H_Homo ) : 17 cas 2,3 %
Total : 388 cas 52 %
Ces cinq groupes sont composés de 388 personnes, âgées de 18 à 83 ans (M = 31 ans ; s = 11,07), dont 193 femmes de 18 à 55 ans (M = 27,2 ; s = 7,45) et 195 hommes de 18 à 83 ans (M = 34,9 ; s = 12,67). On remarque surtout que les hommes sont plus âgés que les femmes (entre 8 à 9 ans de plus en moyenne). Les principales analyses statistiques présentées dans ce document portent sur les 4 premiers groupes. Le groupe des hommes homosexuels n'étant constitué que de 17 cas, le résultat de leur analyse est sujet à cautions. Les groupes homo-, bi- et hétéro-sexuels ont été constitués en fonction de l'activité sexuelle des participants.
Plusieurs rubriques du questionnaire étaient spécialement destinées à recueillir des informations générales sur les participants et sur leurs activités sexuelles (questions n°2, 3, 6, 7, 9, 11, 14, 15, 20, 21 & 22). Les données récoltées ont permis de préciser le profil général de la sexualité des participants à l'enquête.
|
Groupe (genre + orientation) |
Âge moyen |
N |
Écart type |
|
H_Hété |
35,68 |
114 |
12,593 |
|
F_Hété |
27,10 |
135 |
7,178 |
|
F_Bi |
( 26,44 ) 27,36 |
58 |
8,124 |
|
H_Bi |
( 34,14 ) 36,85 |
64 |
12,589 |
|
H_Homo |
23,00 |
17 |
5,601 |
|
Total |
31,03 |
388 |
11,066 |
Remarque : les nombres en italique correspondent aux personnes les plus bisexuelles ( 3, 4 et 5 sur l'échelle de Kinsey )
Pour la grande majorité des personnes, l'entrée dans la sexualité a été progressive : d'abord des activités autoérotiques (masturbation), puis des activités érotiques non coïtales (caresses sensuelles, baiser, masturbation réciproque, etc.), puis le coït vaginal, et enfin, pour certains, le coït anal. Cette initiation sexuelle s'étend presque sur une dizaine d'année, entre les âges de 10 à 20 ans.
Chaque groupe d'orientation sexuelle (hétérosexuel, bisexuel et homosexuel) a un profil d'activités érotiques distincts des autres.
La plupart des hétérosexuels ont eu entre 1 à 10 partenaires tandis que la majorité des homosexuels et surtout des bisexuels ont eu entre 5 à 50 partenaires. Un tiers des hommes bisexuels ont eu en moyenne entre 20 à 50 partenaires. La fréquence moyenne des activité sexuelles avec un partenaire est de plusieurs fois par semaine. La fréquence des homosexuels et un peu plus faible tandis que celle des bisexuels est un peu plus importante. Environ 20% des personnes les plus bisexuelles (3, 4 et 5 sur l'échelle de Kinsey) ont des activités érotiques plusieurs fois par jour. La quasi totalité (environ 90%) des hétérosexuels ont actuellement un partenaire et en sont amoureux. Les bisexuels, et surtout les femmes bisexuelles, sont près d'un tiers à ne pas avoir actuellement de partenaire et un tiers à ne pas être amoureuse de leur partenaire actuel.
Les hétérosexuels ne pratiquent quasiment pas l'échangisme ou la sexualité en groupe, mais environ 5 à 10% d'entre eux ont essayé des pratiques telles le triolisme, l'ondinisme ou le sado-masochisme, et 2 à 3 % les pratiquent régulièrement. Les bisexuels, et surtout les personnes les plus bisexuelles, sont plus nombreux (entre 20 à 30%) à avoir essayé ces pratiques plusieurs fois, en particulier le triolisme, et environ 5 à 15 % les pratiquent régulièrement. Les femmes les plus bisexuelles ont la sexualité la plus diversifiée.
Quand elles n'ont pas de partenaires, la majorité des personnes ont des activités autoérotiques en moyenne une fois par semaine. Les femmes se masturbent moins (environ une fois par semaine) que les hommes tandis que les hommes les plus bisexuel se masturbent de une à plusieurs fois par jour.
Par rapport aux désirs et aux fantasmes sexuels, les bisexuels ont plus de fantasmes que les hétérosexuels et que le contenu de ces fantasmes correspond à une sexualité plus diversifiée. À noter que pour tous les groupes les fantasmes sexuels sont plus débridés que la sexualité réellement vécue, mais correspondent globalement à l'orientation sexuelle des activités érotiques réellement pratiquées.
Le plaisir sexuel est important ou très important pour la quasi totalité des personnes, tandis que paradoxalement l'orgasme est jugé relativement moins important que le plaisir érotique. La sexualité occupe une grande importance dans la vie de la majorité des participants à l'enquête. Les bisexuels, et surtout les hommes bisexuels, accordent plus que les autres une plus grande importance à tous ces aspects de la sexualité.
Dans chaque groupe d'orientation sexuelle, les différences entre le profil des femmes et celui des hommes sont faibles. Les différences hommes / femmes sont moins importantes que les différences hétérosexuel / bisexuel / homosexuel.
En conclusion, les femmes et les hommes ont un profil sexuel relativement similaire et c'est au niveau de l'orientation des activités sexuelles qu'on observe des différences plus marquées. Les homosexuels et les bisexuels, et surtout les personnes les plus bisexuelles (uniquement 3, 4 et 5 sur l'échelle de Kinsey), ont la sexualité la plus précoce, la plus intense et la plus diversifiée.
En raison de ces profils distincts, les différents groupes d'orientation sexuelle n'ont pas été regroupés en un seul pour les analyses statistiques et la vérification des hypothèses.
Les informations détaillées relatives aux participants, au profil des activités sexuelles, aux critères de sélection et de constitution des groupes sont données en annexe, à la section "Critères de sélection des participants & Formations des groupes", page 179.
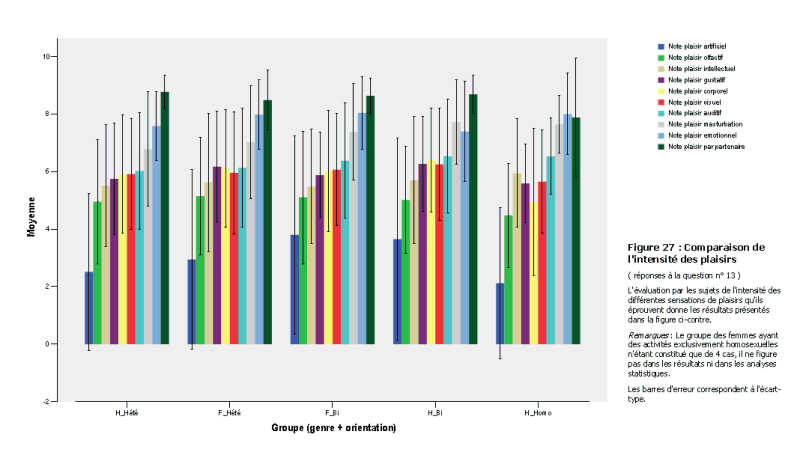
 |
4 – Résultats
a – Le plaisir érotique est-il un des plaisirs les plus intenses que l'organisme peut éprouver ?
L'hypothèse intermédiaire Hy1 , "le plaisir érotique est un des plaisirs les plus intenses que l'organisme peut éprouver", est testée en utilisant deux questions : une, principale, spécifique à l'intensité du plaisir perçu, l'autre, complémentaire, spécifique aux activités préférées.
( questions n° 13 & 1, en pages 6 et 2 du questionnaire présenté à l'annexe II )
a – Comparaison des différentes sensations de plaisir
( réponses à la question n° 13 )
Quel que soit le groupe, on observe un profil relativement similaire de l'intensité perçue des différents types de plaisir. Le plaisir érotique provoqué par un partenaire est en première position, immédiatement suivit par le plaisir émotionnel et le plaisir par masturbation (Figure 27). A priori, il semble bien que le plaisir érotique soit un des plaisirs les plus intenses que l'organisme peut éprouver.
Les analyses statistiques, réalisées dans les sections suivantes, permettront de vérifier si ces résultats sont significatifs.
Groupe : Hommes hétérosexuels
Les résultats détaillées des réponses du groupe des 114 hommes hétérosexuels sont donnés dans le tableau ci-dessous :
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Plaisir sexuel par partenaire |
8,77 |
,585 |
112 |
5 |
9 |
|
Plaisir émotionnel |
7,58 |
1,193 |
64 |
5 |
9 |
|
Plaisir sexuel par masturbation |
6,78 |
1,984 |
111 |
1 |
9 |
|
Plaisir auditif |
6,02 |
2,027 |
111 |
0 |
9 |
|
Plaisir corporel |
5,91 |
2,047 |
111 |
0 |
9 |
|
Plaisir visuel |
5,91 |
1,928 |
111 |
1 |
9 |
|
Plaisir gustatif |
5,75 |
1,938 |
111 |
0 |
9 |
|
Plaisir intellectuel |
5,50 |
2,123 |
111 |
0 |
9 |
|
Plaisir olfactif |
4,95 |
2,151 |
111 |
0 |
9 |
|
Plaisir artificiel (drogue, etc.) |
2,51 |
2,727 |
109 |
0 |
9 |
Tableau 1 : Classement de l'intensité des plaisirs des hommes hétérosexuels
Dans un premier temps, on cherche à savoir s'il existe des différences significatives entre les différents plaisirs, au moyen d'un test statistique de comparaison de plusieurs moyennes (test de Friedman [6] ). Le résultat du test ( n = 61 ; Khi-deux = 248,38 ; ddl = 9 ; p < 0,001 ) indique qu'il existe une différence statistiquement significative entre les moyennes des notes données aux différents types de plaisir.
Ensuite, on cherche à savoir si les plaisirs érotiques sont supérieurs aux autres plaisirs. Comme l'hypothèse Hy1 – le plaisir érotique est un des plaisirs les plus intenses que l'organisme peut éprouver – est une hypothèse a priori, les moyennes des plaisirs érotiques sont simplement comparés deux à deux avec les moyennes des autres types de plaisir, en utilisant un test approprié (test de Wilcoxon [7] ). Le seuil de significativité est ici fixé à 0,01, uniquement pour ces comparaisons multiples. On obtient les résultats suivants :
|
|
Note plaisir émotionnel - Note plaisir par partenaire |
Note plaisir émotionnel - Note plaisir masturbation |
Note plaisir auditif - Note plaisir masturbation |
Note plaisir corporel - Note plaisir masturbation |
Note plaisir visuel - Note plaisir masturbation |
Note plaisir intellectuel - Note plaisir masturbation |
|
Z |
-5,414(a) |
-2,227(b) |
-3,111(a) |
-3,473(a) |
-3,464(a) |
-4,530(a) |
|
Asymp. Sig. (2-tailed) |
,000 |
,026 |
,002 |
,001 |
,001 |
,000 |
( NB : présentation des comparaisons des moyennes les plus proches ; les autres comparaisons sont significatives à 0,001. )
On observe que la moyenne des notes données au plaisir érotique procuré par un partenaire est statistiquement différente de toutes les autres moyennes. Quant au plaisir érotique procuré par soi-même, sa moyenne est statistiquement différente de toutes les autres moyennes, sauf de celle du plaisir émotionnel. Il apparaît donc trois groupes distincts d'intensité de plaisirs : 1) le plaisir érotique procuré par un partenaire ; 2) le plaisir émotionnel et le plaisir érotique procuré par soi-même ; et 3) tous les autres plaisirs.
D'où le plaisir érotique procuré par un partenaire est le plaisir le plus intense que ces personnes peuvent éprouver, tandis que le plaisir érotique procuré par soi-même n'est qu'un des plaisirs les plus intenses que ces personnes peuvent éprouver.
En conclusion, chez les hommes ayant des activités érotiques hétérosexuelles, l'hypothèse intermédiaire Hy1 est donc vérifiée.
Résultats supplémentaires
En complément des analyses précédentes, afin de rechercher d'autres résultats significatifs concernant l'intensité des plaisirs, toutes les différentes moyennes sont comparées 2 à 2 avec un test approprié (analyse post-hoc exploratoire, test de Tukey [8] ). Cette analyse exploratoire (non présentée dans ce document) indique que l'intensité du plaisir artificiel (provoqué par des drogues, des médicaments, etc.) est statistiquement inférieure à tous les autres plaisirs, sauf au plaisir olfactif.
En conclusion de ces analyses statistiques, on observe apparemment 4 groupes d'intensité de plaisir, distincts les uns des autres. Dans l'ordre : 1) le plaisir procuré par un partenaire ; 2) le plaisir émotionnel et le plaisir procuré par soi-même ; 3) les plaisirs auditif, visuel, olfactif, gustatif, corporel et intellectuel ; et peut être, 4) le plaisir procuré artificiellement.
Des analyses complémentaires, réalisées dans les sections suivantes, permettront d'affiner ces résultats.
Groupe : Femmes hétérosexuelles
Les résultats détaillées des réponses du groupe des 136 femmes hétérosexuelles sont donnés dans le tableau ci-dessous :
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Plaisir sexuel par partenaire |
8,49 |
1,046 |
134 |
2 |
9 |
|
Plaisir émotionnel |
7,98 |
1,202 |
46 |
3 |
9 |
|
Plaisir sexuel par masturbation |
7,02 |
1,972 |
135 |
0 |
9 |
|
Plaisir gustatif |
6,17 |
1,921 |
134 |
0 |
9 |
|
Plaisir auditif |
6,13 |
2,065 |
135 |
0 |
9 |
|
Plaisir corporel |
6,10 |
2,035 |
134 |
0 |
9 |
|
Plaisir visuel |
5,96 |
2,127 |
134 |
0 |
9 |
|
Plaisir intellectuel |
5,62 |
2,409 |
135 |
0 |
9 |
|
Plaisir olfactif |
5,15 |
2,035 |
131 |
0 |
9 |
|
Plaisir artificiel (drogue, etc.) |
2,95 |
3,132 |
133 |
0 |
9 |
Tableau 2 : Classement de l'intensité des plaisirs des femmes hétérosexuelles
L'analyse des moyennes (test de Friedman : n = 43 ; Khi-deux = 157,56 ; ddl = 9 ; p < 0,001 ) indique qu'il existe une différence statistiquement significative entre les moyennes des différents types de plaisir.
La comparaison des plaisirs érotiques avec les autres types de plaisir, au seuil de significativité de 0,01, donne les résultats suivants (test de Wilcoxon) :
|
|
Note plaisir émotionnel - Note plaisir par partenaire |
Note plaisir émotionnel - Note plaisir masturbation |
Note plaisir gustatif - Note plaisir masturbation |
Note plaisir auditif - Note plaisir masturbation |
Note plaisir corporel - Note plaisir masturbation |
Note plaisir intellectuel - Note plaisir masturbation |
|
Z |
-2,238(a) |
-3,213(b) |
-4,116(a) |
-3,576(a) |
-4,339(a) |
-4,876(a) |
|
Asymp. Sig. (2-tailed) |
,025 |
,001 |
,000 |
,000 |
,000 |
,000 |
( NB : présentation des comparaisons des moyennes les plus proches ; les autres comparaisons sont significatives à 0,001. )
On observe que la moyenne des notes données au plaisir érotique procuré par un partenaire est statistiquement différente de toutes les autres moyennes, sauf de celle du plaisir émotionnel. Quant au plaisir érotique procuré par soi-même, sa moyenne est statistiquement différente de tous les autres moyennes. Il apparaît donc également trois groupes distincts d'intensité de plaisirs : 1) le plaisir érotique procuré par un partenaire et le plaisir émotionnel ; 2) le plaisir érotique procuré par soi-même ; et 3) tous les autres plaisirs.
D'où le plaisir érotique procuré par un partenaire et le plaisir émotionnel sont les plaisirs les plus intenses que ces personnes peuvent éprouver, tandis que le plaisir érotique procuré par soi-même n'est qu'un des plaisirs les plus intenses que ces personnes peuvent éprouver.
En conclusion, chez les femmes ayant des activités érotiques hétérosexuelles, l'hypothèse intermédiaire Hy1 est vérifiée.
En complément, une analyse post-hoc exploratoire (test de Tukey), indique, à trois résultats négatifs près (plaisir artificiel VS plaisir olfactif, intellectuel et corporel – mais significatifs avec une simple comparaison 2 à 2), que l'intensité du plaisir artificiel tend à être statistiquement inférieure aux autres plaisir.
En conclusion de ces analyses statistiques, on observe apparemment 3 groupes d'intensité de plaisir, distincts les uns des autres. Dans l'ordre : 1) le plaisir procuré par un partenaire et le plaisir émotionnel ; 2) le plaisir procuré par soi-même ; et 3) les plaisirs auditif, visuel, olfactif, gustatif, corporel, intellectuel et artificiel.
Des analyses complémentaires, réalisées dans les sections suivantes, permettront d'affiner ces résultats.
Groupe : Femmes bisexuelles
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Plaisir sexuel par partenaire |
8,63 |
,616 |
57 |
7 |
9 |
|
Plaisir émotionnel |
8,04 |
1,261 |
23 |
5 |
9 |
|
Plaisir sexuel par masturbation |
7,38 |
1,674 |
58 |
3 |
9 |
|
Plaisir auditif |
6,38 |
2,007 |
58 |
1 |
9 |
|
Plaisir visuel |
6,07 |
1,945 |
58 |
1 |
9 |
|
Plaisir corporel |
6,02 |
2,107 |
58 |
1 |
9 |
|
Plaisir gustatif |
5,88 |
1,499 |
58 |
2 |
9 |
|
Plaisir intellectuel |
5,48 |
1,985 |
58 |
0 |
9 |
|
Plaisir olfactif |
5,10 |
2,299 |
58 |
0 |
9 |
|
Plaisir artificiel (drogue, etc.) |
3,80 |
3,445 |
56 |
0 |
9 |
Tableau 3 : Classement de l'intensité des plaisirs des 58 femmes bisexuelles
L'analyse des moyennes (test de Friedman : n = 22 ; Khi-deux = 96,45 ; ddl = 9 ; p < 0,001) indique qu'il existe une différence statistiquement significative entre les moyennes des différents types de plaisir.
La comparaison des plaisirs érotiques avec les autres types de plaisir, au seuil de significativité de 0,01, donne les résultats suivants (test de Wilcoxon) :
|
|
Note plaisir par partenaire - Note plaisir masturbation |
Note plaisir émotionnel - Note plaisir par partenaire |
Note plaisir émotionnel - Note plaisir masturbation |
Note plaisir auditif - Note plaisir masturbation |
Note plaisir visuel - Note plaisir masturbation |
Note plaisir corporel - Note plaisir masturbation |
|
Z |
-4,474(a) |
-2,223(b) |
-,235(a) |
-3,326(b) |
-4,001(b) |
-3,815(b) |
|
Asymp. Sig. (2-tailed) |
,000 |
,026 |
,814 |
,001 |
,000 |
,000 |
( NB : présentation des comparaisons des moyennes les plus proches ; les autres comparaisons sont significatives à 0,001. )
On observe que la moyenne des notes données au plaisir érotique procuré par un partenaire est statistiquement différente de toutes les autres moyennes, sauf de celle du plaisir émotionnel. Le plaisir érotique procuré par soi-même est significativement distinct de tous les autres plaisirs, sauf des plaisirs émotionnel et érotique par partenaire. Néanmoins, ces résultats négatifs sont peut être dû au faible nombre de notes du plaisir émotionnel (22 notes seulement), car autrement le profil des moyennes est similaire entre les femmes bisexuelles et hétérosexuelles. Il apparaît donc trois groupes distincts d'intensité de plaisirs : 1) le plaisir érotique procuré par un partenaire; 2) le plaisir érotique procuré par soi-même, avec le plaisir émotionnel qui n'est pas distinct de ces deux premiers groupe ; et 3) tous les autres plaisirs.
On observe que le plaisir érotique procuré par un partenaire, le plaisir émotionnel et plaisir érotique procuré par soi-même sont les plaisirs les plus intenses que ces personnes peuvent éprouver. Chez les femmes ayant des activités érotiques bisexuelles, l'hypothèse intermédiaire Hy1 est vérifiée.
L'analyse post-hoc exploratoire (Tukey) n'a pas mis en évidence d'autres résultats statistiquement significatifs, vraisemblablement en raison de la petite taille de l'échantillon.
En conclusion, on observe que le groupe des femmes ayant des activités bisexuelles est relativement similaire à celui des hétérosexuels.
Groupe : Hommes bisexuels
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Plaisir sexuel par partenaire |
8,69 |
,664 |
64 |
6 |
9 |
|
Plaisir sexuel par masturbation |
7,72 |
1,474 |
64 |
3 |
9 |
|
Plaisir émotionnel |
7,39 |
1,745 |
36 |
2 |
9 |
|
Plaisir auditif |
6,53 |
1,976 |
64 |
2 |
9 |
|
Plaisir corporel |
6,39 |
1,805 |
64 |
1 |
9 |
|
Plaisir gustatif |
6,27 |
1,648 |
63 |
2 |
9 |
|
Plaisir visuel |
6,25 |
1,944 |
64 |
2 |
9 |
|
Plaisir intellectuel |
5,70 |
2,216 |
64 |
1 |
9 |
|
Plaisir olfactif |
5,02 |
1,856 |
64 |
0 |
9 |
|
Plaisir artificiel (drogue, etc.) |
3,66 |
3,506 |
64 |
0 |
9 |
Tableau 4 : Classement de l'intensité des plaisirs des 64 hommes bisexuels
L'analyse des moyennes (test de Friedman : n = 16 ; Khi-deux = 73,33 ; dll = 9 ; p < 0,001) indique qu'il existe une différence statistiquement significative entre les moyennes des différents types de plaisir.
La comparaison des plaisirs érotiques avec les autres types de plaisir, au seuil de significativité de 0,01, donne les résultats suivants (test de Wilcoxon) :
|
|
Note plaisir par partenaire - Note plaisir masturbation |
Note plaisir émotionnel - Note plaisir par partenaire |
Note plaisir émotionnel - Note plaisir masturbation |
Note plaisir auditif - Note plaisir masturbation |
Note plaisir visuel - Note plaisir masturbation |
|
Z |
-4,090(a) |
-3,894(b) |
-1,487(b) |
-3,345(b) |
-4,307(b) |
|
Asymp. Sig. (2-tailed) |
,000 |
,000 |
,137 |
,001 |
,000 |
( NB : présentation des comparaisons des moyennes les plus proches ; les autres comparaisons sont significatives à 0,001. )
On observe que le plaisir érotique procuré par un partenaire est le plaisir le plus intense que ces personnes peuvent éprouver, tandis que le plaisir érotique procuré par soi-même n'est qu'un des plaisirs les plus intenses qu'elles peuvent éprouver. Chez les hommes ayant des activités érotiques bisexuelles, l'hypothèse intermédiaire Hy1 est donc vérifiée.
L'analyse post-hoc exploratoire (Tukey) n'a pas mis en évidence d'autres résultats statistiquement significatifs, vraisemblablement en raison de la petite taille de l'échantillon.
En conclusion, on observe que le profil du groupe des hommes ayant des activités bisexuelles est très similaire à celui des hétérosexuels.
Groupe : Hommes homosexuels
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Plaisir émotionnel |
9,00 |
. |
(un seul cas) 1 |
9 |
9 |
|
Plaisir sexuel par partenaire |
7,88 |
2,125 |
16 |
1 |
9 |
|
Plaisir sexuel par masturbation |
7,56 |
,964 |
16 |
6 |
9 |
|
Plaisir auditif |
6,44 |
1,315 |
16 |
4 |
9 |
|
Plaisir intellectuel |
6,06 |
1,879 |
16 |
2 |
9 |
|
Plaisir visuel |
5,69 |
1,852 |
16 |
3 |
9 |
|
Plaisir gustatif |
5,44 |
1,263 |
16 |
4 |
8 |
|
Plaisir corporel |
5,06 |
2,594 |
16 |
1 |
9 |
|
Plaisir olfactif |
4,38 |
1,821 |
16 |
2 |
8 |
|
Plaisir artificiel (drogue, etc.) |
2,25 |
2,646 |
16 |
0 |
9 |
Tableau 5 : Classement de l'intensité des plaisirs des 17 hommes homosexuels
L'analyse des moyennes (test de Friedman : n = 16 ; Khi-deux = 58,98 ; dll = 8 ; p < 0,001) indique qu'il existe une différence statistiquement significative entre les moyennes des différents type de plaisir.
La comparaison des plaisirs érotiques avec les autres types de plaisir, au seuil de significativité de 0,01, donne les résultats suivants (test de Wilcoxon) :
|
|
Note plaisir par partenaire - Note plaisir masturbation |
Note plaisir auditif - Note plaisir par partenaire |
Note plaisir intellectuel - Note plaisir par partenaire |
Note plaisir visuel - Note plaisir par partenaire |
Note plaisir corporel - Note plaisir par partenaire |
Note plaisir gustatif - Note plaisir par partenaire |
|
Z |
-1,134(a) |
-2,176(b) |
-2,370(b) |
-2,351(b) |
-2,864(b) |
-2,838(b) |
|
Asymp. Sig. (2-tailed) |
,257 |
,030 |
,018 |
,019 |
,004 |
,005 |
|
|
Note plaisir auditif - Note plaisir masturbation |
Note plaisir intellectuel - Note plaisir masturbation |
Note plaisir visuel - Note plaisir masturbation |
Note plaisir corporel - Note plaisir masturbation |
Note plaisir gustatif - Note plaisir masturbation |
Note plaisir olfactif - Note plaisir masturbation |
|
Z |
-2,403(a) |
-2,220(a) |
-3,109(a) |
-2,772(a) |
-3,424(a) |
-3,372(a) |
|
Asymp. Sig. (2-tailed) |
,016 |
,026 |
,002 |
,006 |
,001 |
,001 |
( NB : présentation des comparaisons des moyennes les plus proches ; les autres comparaisons sont significatives à 0,001. )
On observe dans ce groupe que les plaisirs érotiques sont distincts ou tendent à être significativement distincts des autres types de plaisirs, mais aucun plaisir érotique n'est statistiquement différent de tous les autres. L'analyse post-hoc exploratoire (Tukey) n'a pas mis en évidence d'autres résultats significatifs. Chez les hommes ayant des activités érotiques homosexuelles, l'hypothèse intermédiaire Hy1 n'est donc pas vérifiée, peut être en raison de la petite taille du groupe.
En conclusion, on n'observe pas de résultats significatifs, mais le profil du groupe des hommes ayant des activités homosexuelles est assez similaire à celui des hétérosexuels.
Conclusion
Au terme des analyses statistiques, on observe chez tous les participants à cette étude – à l'exclusion du petit groupe des homosexuels – apparemment 3 groupes d'intensité de plaisir, distincts les uns des autres. Dans l'ordre : 1) le plaisir érotique procuré par un partenaire; 2) le plaisir érotique procuré par soi-même, avec le plaisir émotionnel qui n'est pas toujours distinct de ces deux premiers groupe ; et 3) tous les autres plaisirs.
Des analyses complémentaires, réalisées dans les sections suivantes, permettront d'affiner ces résultats.
En conclusion, quel que soit le groupe, l'hypothèse intermédiaire Hy1 est toujours vérifiée.
a.1 – Influence des souvenirs dans l'intensité du plaisir sexuel par auto-stimulation
( réponses à la question n° 4, en pages 3 du questionnaire présenté à l'annexe II )
Les analyses statistiques précédemment réalisées ont montré que le plaisir érotique procuré par un partenaire est significativement supérieur au plaisir érotique provoqué par soi-même. Par ailleurs ont sait que la masturbation est rarement effectuée juste par la simple stimulation du pénis/clitoris, et qu'elle s'accompagne souvent de différents supports externes et/ou d'évocations psychiques internes.
D'où il est possible qu'une partie importante de l'intensité du plaisir érotique provoqué par auto-stimulation provienne du rappel mnésique des sensations perçues au cours des activités sexuelles avec des partenaires.
Une rubrique du questionnaire a été spécifiquement élaborée afin d'apporter des précisions quand aux sources de l'excitation érotique des activités masturbatoires. La forme finale de cette question n'était présente que dans la dernière version du questionnaire, d'où le nombre de réponses est un peu plus réduit (205 réponses données par 118 hommes et 87 femmes). Par ailleurs, comme cette analyse est uniquement complémentaire et que les réponses données à cette question n'ont pas été influencées par l'orientation des activités érotiques, les groupes hétéro-, homo- et bi-sexuels n'ont pas été distingués. Les résultats sont présentés ci-dessous :
Groupe : Femmes
Les résultats détaillées des réponses des 87 femmes sont donnés dans le tableau ci-dessous :
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Source excitation - Fantasmes |
7,92 |
1,422 |
72 |
4 |
9 |
|
Source excitation - Vécu |
7,23 |
1,820 |
61 |
2 |
9 |
|
Combinaison de plusieurs sources |
6,84 |
2,200 |
38 |
1 |
9 |
|
Source excitation - Vidéo |
6,57 |
2,291 |
51 |
1 |
9 |
|
Source excitation - Lecture |
5,98 |
1,928 |
40 |
2 |
9 |
|
Souvenirs de scènes sexuelles |
5,95 |
2,428 |
40 |
1 |
9 |
|
Source excitation - Image |
5,87 |
2,029 |
38 |
2 |
9 |
|
Uniquement la stimulation du corps |
5,79 |
2,397 |
34 |
1 |
9 |
|
Discussion par Internet, téléphone … |
5,23 |
2,208 |
30 |
0 |
9 |
|
Observation (discrète) de personnes |
2,92 |
2,290 |
13 |
0 |
7 |
Tableau 6 : Classement des sources de l'excitation auto-érotique des femmes
Groupe : Hommes
Les résultats détaillées des réponses des 118 hommes sont donnés dans le tableau ci-dessous :
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Source excitation - Vidéo |
7,41 |
1,903 |
97 |
1 |
9 |
|
Combinaison de plusieurs sources |
7,39 |
1,801 |
61 |
1 |
9 |
|
Source excitation - Vécu |
7,30 |
1,690 |
82 |
1 |
9 |
|
Source excitation - Fantasmes |
7,27 |
1,866 |
96 |
1 |
9 |
|
Source excitation - Image |
6,05 |
2,024 |
81 |
0 |
9 |
|
Source excitation - Lecture |
5,98 |
2,128 |
56 |
1 |
9 |
|
Souvenirs de scènes sexuelles |
5,95 |
2,028 |
61 |
1 |
9 |
|
Discussion par Internet, téléphone … |
5,87 |
2,526 |
31 |
0 |
9 |
|
Observation (discrète) de personnes |
5,33 |
2,614 |
21 |
1 |
9 |
|
Uniquement la stimulation du corps |
4,77 |
2,429 |
22 |
1 |
9 |
Tableau 7 : Classement des sources de l'excitation auto-érotique des hommes
On observe, tant chez les femmes que chez les hommes, que l'autostimulation est quasi systématiquement accompagnée de souvenirs de vécu sexuel, de fantasmes et de vidéos érotiques, le plus souvent simultanément combinés durant l'activité masturbatoire. Les deux groupes ont un profil très similaire, et les principales sources de l'excitation érotique sont des supports externes et/ou internes d'activités érotiques avec des partenaires
On peut conclure que la source de l'excitation sexuelle au cours de la masturbation ne se limite pas uniquement à l'autostimulation du corps (uniquement 1 cas sur les 205 réponses), mais s'accompagne presque toujours de différentes activités dont le point commun est vraisemblablement le rappel mnésique des sensations perçues au cours d'activités sexuelles antérieures, vécues avec des partenaires.
Les données recueillies ne permettent pas d'évaluer, dans l'intensité globale du plaisir masturbatoire, les parts respectives du plaisir provoqué uniquement par l'autostimulation du corps de celle provoqué par le rappel mnésique. Néanmoins ces données suggèrent que l'intensité du plaisir sexuel provoqué uniquement par autostimulation, sans aucune activité psychique, est très certainement inférieure à celle recueillie par ce questionnaire (elle pourrait peut être se situer au niveau du plaisir corporel). Le plaisir sexuel provoqué par un partenaire serait donc, de loin, le plaisir érotique le plus intense que l'organisme peut éprouver.
a.2 – Existe-t-il une différence entre les hommes et les femmes par rapport à l'intensité des différents plaisirs ?
Une hypothèse complémentaire à l'hypothèse principale testée dans cette enquête, est que les réactions hédoniques de base seraient indépendantes du genre et seraient similaires chez les femmes et chez les hommes.
L'exploration graphiques des données semble confirmer cette hypothèse.
|
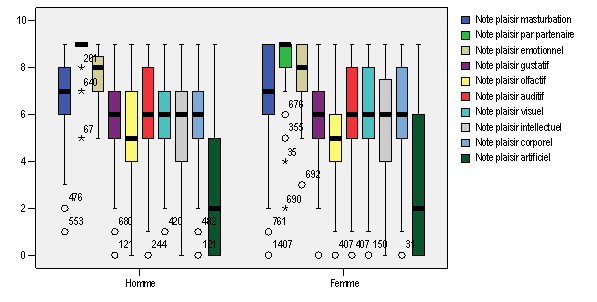
Afin de vérifier si les données des groupes "Homme" et "Femme" sont identiques, les moyennes des données sont comparées pour chaque variable afin de vérifier s'il existe ou non une différence statistiquement significative entre ces deux groupes.
Les résultats de l'analyse statistique (Mann-Whitney [9] ) sont donnés dans le tableau ci-dessous :
|
|
Note plaisir masturbation |
Note plaisir par partenaire |
Note plaisir emotionnel |
Note plaisir gustatif |
Note plaisir olfactif |
Note plaisir auditif |
Note plaisir visuel |
Note plaisir intellectuel |
Note plaisir corporel |
Note plaisir artificiel |
|
Mann-Whitney U |
6903,500 |
6484,000 |
1158,000 |
6504,000 |
6945,500 |
7272,000 |
7308,000 |
7094,500 |
7043,500 |
6897,500 |
|
Wilcoxon W |
13119,500 |
15529,000 |
3238,000 |
12720,000 |
13161,500 |
13488,000 |
13524,000 |
13310,500 |
13259,500 |
12892,500 |
|
Z |
-1,082 |
-2,432 |
-1,983 |
-1,717 |
-,605 |
-,402 |
-,236 |
-,724 |
-,721 |
-,672 |
|
Asymp. Sig. (2-tailed) |
,279 |
,015 |
,047 |
,086 |
,545 |
,688 |
,813 |
,469 |
,471 |
,501 |
On observe qu'il existe une différence statistiquement significative pour les items "plaisir par partenaire" et "plaisir émotionnel", et que l'item "plaisir gustatif" tend à être significativement différent. Les groupes "Homme" et "Femme" ne sont donc pas identiques. L'intensité du plaisir érotique est plus forte chez les hommes tandis que l'intensité du plaisir émotionnel est plus grande chez les femmes.
Néanmoins, on observe, sur la visualisation graphique et sur les tableaux des moyennes, une relative similitude des données entre les deux groupes.
En conclusion, il ne semble pas exister beaucoup de différences entre les hommes et les femmes par rapport à l'intensité des différents types de plaisirs. Au niveau neurophysiologique, il semble donc exister, indépendamment du genre, non pas une identité, mais une similitude des réactions hédoniques.
a.3 – Analyse factorielle exploratoire
Une analyse en composantes principales (ACP), que ce soit sur l'ensemble des réponses ou sur les données de chaque groupe, met en évidence deux facteurs de valeur propre supérieure à 1. La majorité des items se projetant fortement sur le premier facteur, nous avons décidé de ne conserver qu'un facteur général. Les résultats ci-dessous proviennent de l'ACP effectuée sur les réponses de tous les participants à la question n° 13, en supprimant de l'analyse les items qui ont des saturations faibles sur le premier facteur, à savoir les items "Plaisir sexuel par masturbation", "Plaisir sexuel procuré par un partenaire" et "Plaisir artificiel". La solution retenue, présentée dans le Tableau 8, est composée de 7 items qui expliquent 51,4 % de la variance totale. La consistance interne de ce facteur est tout à fait satisfaisante (? de Cronbach = 0,84 ) : les 7 items sont homogènes entre-eux.
|
Item |
Facteur principal |
|
Plaisir gustatif |
,714 |
|
Plaisir olfactif |
,775 |
|
Plaisir auditif |
,818 |
|
Note plaisir visuel |
,775 |
|
Plaisir intellectuel |
,653 |
|
Plaisir corporel |
,579 |
|
Plaisir émotionnel |
,673 |
Tableau 8 : Résultats de l'ACP effectuée sur les réponses à la question n° 13, en regroupant tous les participants ( N = 168 ).
Nous interprétons le facteur principal de la manière suivante : ce facteur correspondrait à une même catégorie ou type de plaisirs, qui seraient engendrés par des processus neurobiologiques identiques ou similaires. A partir des résultats de cette analyse factorielle, et en excluant le plaisir artificiel car provoqué par des drogues ou des substances pharmacologiques, il semblerait exister deux catégories bien distinctes de plaisirs naturels : les plaisirs sexuels et les plaisirs non sexuels. Comme nous avons vu que le plaisir procuré par la masturbation semble provenir en partie du rappel mnésique du plaisir sexuel vécu antérieurement avec un partenaire, ce plaisir érotique procuré par un partenaire apparaît comme étant un plaisir bien distinct, de nature différente et plus intense que tous les autres plaisirs.
a 4 – Analyses complémentaires
Deux analyses complémentaires, relatives aux plaisirs érotique, émotionnel et artificiel, sont effectuées afin d'obtenir des précisions supplémentaires.
Remarque : Afin de limiter la taille de ce document, ne sont donnés ci-dessous que les résultats finaux de ces analyses complémentaires.
Plaisir émotionnel et érotique
Afin d'évaluer la supériorité de l'intensité du plaisir érotique procuré par un partenaire sur l'intensité du plaisir émotionnel, les personnes percevant les plus grandes intensités de plaisir émotionnel ont été regroupées pour une analyse complémentaire.
En regroupant toutes les réponses (110 cas) où le plaisir émotionnel est maximal (notes égales à 8 et 9), on observe que l'intensité perçue du plaisir émotionnel tend à être significativement inférieure à l'intensité du plaisir érotique avec un partenaire (érotique = 8,65 et émotionnel = 8,55, avec p = 0,086). Ce résultat suggère que le plaisir érotique provoqué par un partenaire est bien le plaisir le plus intense que ces personnes peuvent éprouver.
Plaisir artificiel
Cette analyse complémentaire a été effectuée afin de savoir si le procuré artificiellement pourrait être dans certains cas plus intense que les plaisirs naturels.
En regroupant toutes les réponses (51 cas) où le plaisir artificiel est maximal (notes égales à 8 et 9), on observe que les plaisirs émotionnel et érotique avec un partenaire sont aussi intenses que le plaisir artificiel. On n'observe pas de différences significative entre les moyennes. Ce résultat suggère que l'intensité du plaisir artificiel dépend de la substance introduite dans l'organisme et qu'elle n'est jamais supérieure à un plaisir "naturel". La substance introduite ne ferait qu'activer artificiellement des structures neurales impliquées dans la genèse des plaisirs émotionnels et érotiques.
a.5 – Conclusion
À l'issue de toutes ces analyses statistiques et complémentaires relatives aux intensités perçues des différents types de plaisir, on observe apparemment 4 groupes de nature et d'intensité de plaisir, qui semblent distincts les uns des autres. Dans l'ordre :
1) le plaisir érotique procuré par un partenaire, qui serait de nature différente aux autres plaisirs naturels ;
2) le plaisir procuré par des émotions (qui dans certains groupes n'est pas statistiquement différent du plaisir érotique par partenaire) ;
3) le plaisir procuré par des processus non émotionnels : les plaisirs auditif, visuel, olfactif, gustatif, intellectuel, corporel et masturbatoire (sans rappel mnésique des souvenirs du plaisir par partenaire) ;
4) le plaisir procuré artificiellement (dont l'intensité dépend de la substance introduite dans l'organisme ; ce plaisir peut être aussi intense que le plaisir érotique procuré par un partenaire).
En conclusion, quel que soit le genre ou l'orientation des activités sexuelles, l'hypothèse intermédiaire Hy1 est toujours vérifiée.
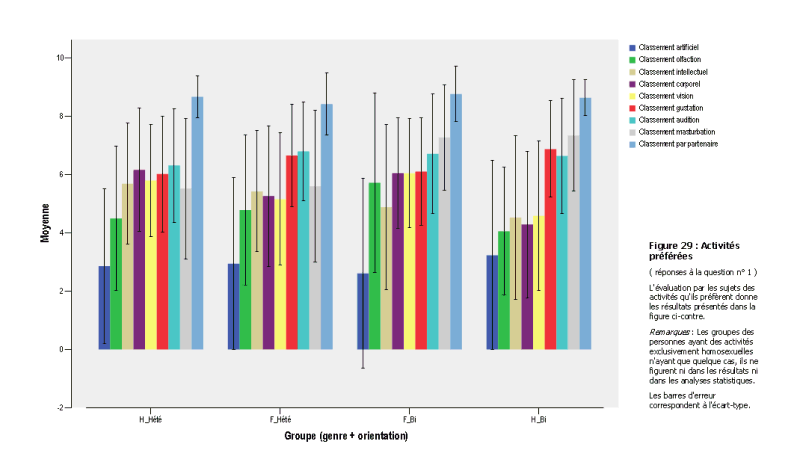
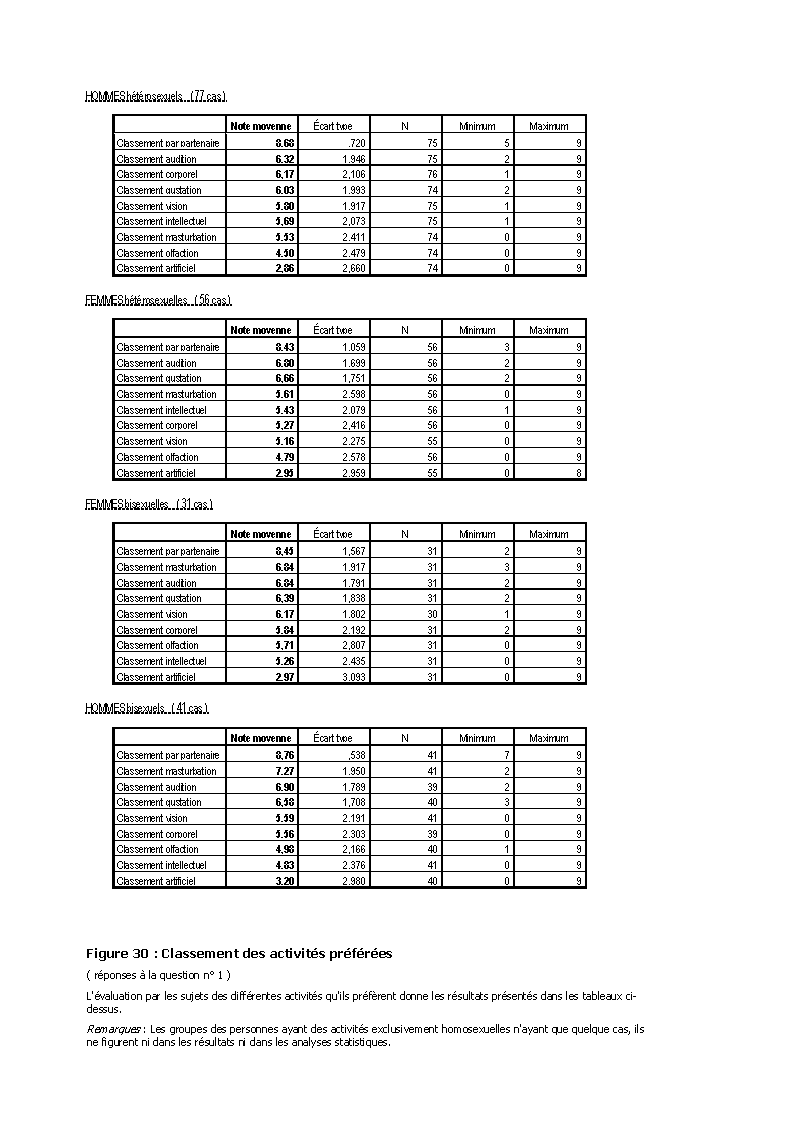
b – Comparaison des activités préférées
( réponses à la question n°1 )
Dans la troisième version du questionnaire, l'évaluation par les sujets (87 femmes et 118 hommes) des différentes activités qu'ils préfèrent indique une nette préférence pour les activités érotiques avec un partenaire (Figure 29 et Figure 30).
Les analyses statistiques confirment, dans tous les groupes, qu'il existe des différences significatives entre les moyennes (p<0,001), et que l'activité sexuelle avec un partenaire est significativement préférée (p<0,001). Les analyses exploratoires post hoc n'indiquent pas d'autres résultats significatifs. Ces résultats mettent en évidence que l'activité érotique avec un partenaire est une activité qui possède des caractéristiques hédoniques et appétitives nettement supérieure à celles des autres activités.
(Voir également en annexe, page 196, les réponses données dans les deux premières versions du questionnaire, avec un autre système de notation.)
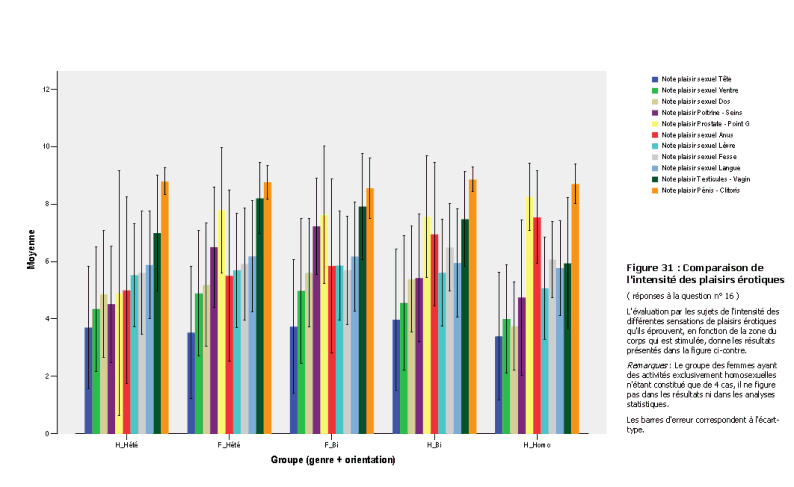
b – Les zones érogènes des organes génitaux sont-elles à l'origine du plaisir érotique le plus intense ?
L'hypothèse intermédiaire Hy2 , "les zones érogènes des organes génitaux sont à l'origine d'un des plaisirs érotiques les plus intenses", est testée en utilisant deux questions : une, principale, spécifique à l'intensité du plaisir érotique perçu, l'autre, complémentaire, spécifique aux activités érotiques préférées.
( questions n° 16 & 17, en pages 7 et 8 du questionnaire présenté à l'annexe II )
a – Comparaison des différentes sensations de plaisir érotique, en fonction des zones érogènes
( réponses à la question n° 16 )
Quel que soit le groupe, on observe que les zones érogènes des organes génitaux (pénis/clitoris, vagin) procurent le plaisir érotique le plus intense (Figure 31).
D'autres zones corporelles procurent également du plaisir érotique. Dans l'ordre chez les femmes : le point G, les seins, la zone orale (lèvre et surtout langue), la zone anale, et, en dernier, le dos, le ventre et la tête. Chez les hommes, l'intensité des autres zones que le pénis semble dépendre de l'orientation des activités érotiques. Dans l'ordre, on trouve les testicules, puis la zone orale pour les hétérosexuels mais la zone anale pour les bisexuels et les homosexuels, puis la zone orale pour les bisexuels et les homosexuels et la zone anale pour les hétérosexuels, et en dernier, comme chez les femmes, le tronc et la tête. En plus des régions mentionnées dans le questionnaire, une minorité de participants (dont surtout 10% des femmes) indique également comme zones érogènes le cou, la nuque et les oreilles, et, indiqués uniquement par quelques personnes, les cuisses, les jambes, les pieds, les bras, la main, les doigts et les paupières.
Les analyses statistiques, réalisées dans les sections suivantes, permettront de vérifier si ces résultats sont significatifs.
Remarques : Principalement en raison de la diversité des pratiques sexuelles individuelles, mais également en raison de caractéristiques neurophysiologiques particulières à chaque zone corporelle, les différentes régions du corps ne sont pas toutes stimulées durant les activités érotiques. Les données recueillies reflètent cette réalité et certaines zones corporelles ne sont pas ou peu citées, et dans certains cas le faible nombre de données disponibles rend les calculs statistiques impossibles ou peu fiables. Certaines données, quand elles sont insuffisantes, ne seront donc pas incluses dans les analyses statistiques.
Groupe : Hommes hétérosexuels
Les résultats détaillées des réponses du groupe des 114 hommes hétérosexuels sont donnés dans le tableau ci-dessous :
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Plaisir sexuel Pénis |
8,79 |
,470 |
114 |
7 |
9 |
|
Plaisir sexuel Testicules |
6,99 |
2,021 |
109 |
1 |
9 |
|
Plaisir sexuel Langue |
5,88 |
1,878 |
101 |
2 |
9 |
|
Plaisir sexuel Fesse |
5,60 |
2,150 |
101 |
0 |
9 |
|
Plaisir sexuel Lèvre |
5,52 |
1,787 |
103 |
2 |
9 |
|
Plaisir sexuel Anus |
5,00 |
3,240 |
90 |
0 |
9 |
|
Plaisir sexuel Prostate |
4,89 |
4,260 |
38 |
0 |
9 |
|
Plaisir sexuel Dos |
4,87 |
2,200 |
92 |
0 |
9 |
|
Plaisir sexuel Poitrine |
4,52 |
2,025 |
93 |
0 |
9 |
|
Plaisir sexuel Ventre |
4,35 |
2,170 |
89 |
0 |
9 |
|
Plaisir sexuel Tête |
3,70 |
2,132 |
91 |
0 |
8 |
Tableau 9 : Classement de l'érogénéité des zones corporelles des hommes hétérosexuels
Dans un premier temps, on cherche à savoir s'il existe des différences significatives entre les différentes zones érogènes, au moyen d'un test statistique de comparaison de plusieurs moyennes (test de Friedman [10] ). Le résultat du test ( n = 30 ; Khi-deux = 113,62 ; ddl = 10 ; p < 0,001 ) indique qu'il existe une différence statistiquement significative entre les moyennes des notes données aux différentes régions érogènes.
Ensuite, on cherche à savoir si les zones érogènes des organes génitaux sont supérieurs aux autres zones érogènes. Comme l'hypothèse Hy1 – les zones érogènes des organes génitaux sont à l'origine d'un des plaisirs érotiques les plus intense – est une hypothèse a priori, la moyenne de la principale zone érogène génitale (le gland du pénis) est simplement comparée deux à deux avec les moyennes des autres zones érogènes, en utilisant un test approprié (test de Wilcoxon [11], avec un seuil de significativité fixé à 0,01, uniquement pour ces comparaisons multiples ). On obtient les résultats suivants :
|
|
Note plaisir sexuel Testicules - Note plaisir sexuel Pénis |
Note plaisir sexuel Pénis - Note plaisir sexuel Langue |
Note plaisir sexuel Fesse - Note plaisir sexuel Pénis |
Note plaisir sexuel Anus - Note plaisir sexuel Pénis |
Note plaisir sexuel Prostate - Note plaisir sexuel Pénis |
|
Z |
-7,397(a) |
-8,503(b) |
-8,446(a) |
-7,597(a) |
-4,168(a) |
|
Asymp. Sig. (2-tailed) |
,000 |
,000 |
,000 |
,000 |
,000 |
( NB : présentation des comparaisons des moyennes les plus proches ; les autres comparaisons sont significatives à 0,001. )
On observe que la moyenne des notes données à la zone érogène du pénis est statistiquement différente de toutes les autres moyennes. On observe donc que le plaisir érotique procuré par la stimulation du pénis est le plaisir érotique le plus intense que ces personnes peuvent éprouver.
En conclusion, chez les hommes ayant des activités érotiques hétérosexuelles, l'hypothèse intermédiaire Hy2 est donc validée.
Résultats supplémentaires
En complément des analyses précédentes, afin de rechercher d'autres résultats significatifs, toutes les différentes moyennes sont comparées 2 à 2 avec un test approprié (analyse post-hoc exploratoire, test de Tukey [12] ). Cette analyse exploratoire (non présentée dans ce document) a mis en évidence quelques autres résultats significatifs, qui suggèrent l'existence de plusieurs zones corporelles différentes les unes des autres par rapport à leur intensité érogène. On observe apparemment, dans l'ordre, une zone génitale (pénis et testicule), puis une zone orale (lèvre et langue), puis une zone anale (fesse, anus, prostate) et enfin le ventre, le dos et la tête. Néanmoins, principalement en raison des différences interindividuelles dans les pratiques sexuelles, certaines zones corporelles n'ont pas été stimulées par tous les participants. D'où, en raison du petit nombre de notes disponibles pour certaines variables ("prostate" par exemple) et du nombre important de variables à comparer, les analyses statistiques exploratoires manques de puissance. Ces particularités expliquent vraisemblablement une des raisons pour lesquelles les analyses statistiques n'ont pas mis en évidence de différences très significatives entre les différentes zones érogènes non génitales.
Des analyses complémentaires, réalisées dans les sections suivantes, permettront d'affiner ces résultats.
Groupe : Femmes hétérosexuelles
Les résultats détaillées des réponses du groupe des 136 femmes hétérosexuelles sont donnés dans le tableau ci-dessous :
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Plaisir sexuel Clitoris |
8,76 |
,578 |
134 |
6 |
9 |
|
Plaisir sexuel Vagin |
8,20 |
1,236 |
133 |
1 |
9 |
|
Plaisir sexuel Point G |
7,79 |
2,189 |
94 |
0 |
9 |
|
Plaisir sexuel Seins |
6,50 |
2,105 |
135 |
0 |
9 |
|
Plaisir sexuel Langue |
6,18 |
1,932 |
121 |
0 |
9 |
|
Plaisir sexuel Fesse |
5,91 |
1,959 |
125 |
0 |
9 |
|
Plaisir sexuel Lèvre |
5,69 |
1,990 |
127 |
0 |
9 |
|
Plaisir sexuel Anus |
5,50 |
2,976 |
107 |
0 |
9 |
|
Plaisir sexuel Dos |
5,19 |
2,146 |
107 |
0 |
9 |
|
Plaisir sexuel Ventre |
4,89 |
2,190 |
110 |
0 |
8 |
|
Plaisir sexuel Tête |
3,53 |
2,294 |
106 |
0 |
8 |
Tableau 10 : Classement de l'érogénéité des zones corporelles des femmes hétérosexuelles
L'analyse des moyennes (test de Friedman : n = 64 ; Khi-deux = 343,35 ; ddl = 10 ; p < 0,001 ) indique qu'il existe une différence statistiquement significative entre les moyennes des différentes zones érogènes.
La comparaison des zones érogènes génitales (clitoris et vagin) entre elles et avec les autres zones érogènes donne les résultats suivants (test de Wilcoxon, avec un seuil de significativité fixé à 0,01, uniquement pour ces comparaisons multiples) :
|
|
Note plaisir sexuel Vagin - Note plaisir sexuel Clitoris |
Note plaisir sexuel Point G - Note plaisir sexuel Clitoris |
Note plaisir sexuel Clitoris - Note plaisir sexuel Seins |
Note plaisir sexuel Point G - Note plaisir sexuel Vagin |
Note plaisir sexuel Vagin - Note plaisir sexuel Seins |
|
Z |
-4,528(a) |
-3,725(a) |
-8,894(b) |
-2,714(a) |
-7,936(b) |
|
Asymp. Sig. (2-tailed) |
,000 |
,000 |
,000 |
,007 |
,000 |
( NB : présentation des comparaisons des moyennes les plus proches ; les autres comparaisons sont significatives à 0,001. )
On observe donc que le plaisir érotique, procuré par la stimulation des zones érogènes des organes génitaux, et en particulier du clitoris, est le plaisir érotique le plus intense que ces personnes peuvent éprouver.
En conclusion, chez les femmes ayant des activités érotiques hétérosexuelles, l'hypothèse intermédiaire Hy2 est donc validée.
En complément, l'analyse post-hoc exploratoire (Tukey) a mis en évidence que l'item "point G" est significativement inférieur aux items "clitoris" et "vagin", et supérieur à tous les autres items. Mais, bien que le profil des réponses soit similaire à celui des hommes hétérosexuels, il n'apparaît quasiment pas de différences statistiques entre les zones érogènes non génitales. Ce résultat signifierait que les zones érogènes non génitales produiraient des intensités équivalentes de plaisir érotique. Néanmoins, ce résultat statistique est difficilement plausible avec les moyennes (par exemple, la tête ne semble pas être une zone très érogène), et, surtout, est en contradiction avec les connaissances provenant de la sexologie.
En conclusion de toutes ces analyses statistiques, on observe donc 4 groupes de zones érogènes, statistiquement différents les uns des autres. Dans l'ordre : 1) Le clitoris ; 2) le vagin ; 3) le point G ; et, 4) les seins, les lèvres, la langue, les fesses, l'anus, le dos, le ventre et la tête.
Des analyses complémentaires, réalisées dans les sections suivantes, permettront d'affiner ces résultats.
Groupe : Femmes bisexuelles
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Plaisir sexuel Clitoris |
8,55 |
1,046 |
58 |
3 |
9 |
|
Plaisir sexuel Vagin |
7,91 |
1,857 |
58 |
0 |
9 |
|
Plaisir sexuel Point G |
7,63 |
2,388 |
46 |
0 |
9 |
|
Plaisir sexuel Seins |
7,22 |
1,676 |
58 |
2 |
9 |
|
Plaisir sexuel Langue |
6,18 |
1,900 |
57 |
2 |
9 |
|
Plaisir sexuel Lèvre |
5,86 |
1,894 |
57 |
2 |
9 |
|
Plaisir sexuel Anus |
5,85 |
3,038 |
52 |
0 |
9 |
|
Plaisir sexuel Fesse |
5,70 |
1,887 |
53 |
2 |
9 |
|
Plaisir sexuel Dos |
5,60 |
1,884 |
50 |
0 |
9 |
|
Plaisir sexuel Ventre |
4,98 |
2,515 |
47 |
0 |
9 |
|
Plaisir sexuel Tête |
3,74 |
2,317 |
42 |
0 |
8 |
Tableau 11 : Classement de l'érogénéité des zones corporelles des 58 femmes bisexuelles
L'analyse des moyennes (test de Friedman : n = 33 ; Khi-deux = 149,59 ; ddl = 10 ; p < 0,001 ) indique qu'il existe une différence statistiquement significative entre les moyennes des différentes zones érogènes.
La comparaison des zones érogènes génitales avec les autres zones érogènes, au seuil de significativité de 0,01, donne les résultats suivants (test de Wilcoxon) :
|
|
Note plaisir sexuel Vagin - Note plaisir sexuel Clitoris |
Note plaisir sexuel Point G - Note plaisir sexuel Clitoris |
Note plaisir sexuel Clitoris - Note plaisir sexuel Seins |
Note plaisir sexuel Point G - Note plaisir sexuel Vagin |
Note plaisir sexuel Vagin - Note plaisir sexuel Seins |
|
Z |
-2,381(a) |
-2,159(a) |
-4,751(b) |
-,700(a) |
-3,168(b) |
|
Asymp. Sig. (2-tailed) |
,010 |
,031 |
,000 |
,484 |
,002 |
( NB : présentation des comparaisons des moyennes les plus proches ; les autres comparaisons sont significatives à 0,001. )
La seule comparaison qui n'est pas significative est entre les items "point G" avec "vagin" et "clitoris". Mais comme le point G, ou plus précisément une zone particulièrement sensible dans le vagin, participe au plaisir vaginal global, ce résultat indique que pour ce groupe il n'y a pas de différence significative entre le clitoris et le vagin . En conclusion, on observe donc que le plaisir érotique procuré par la stimulation du clitoris ou du vagin est le plaisir érotique le plus intense que ces personnes peuvent éprouver. Chez les femmes ayant des activités érotiques bisexuelles, l'hypothèse intermédiaire Hy2 est donc validée.
L'analyse post-hoc exploratoire (Tukey) n'a pas mis en évidence d'autres résultats significatifs. Néanmoins, l'effectif du groupe étant plus faible que celui des hétérosexuels, les similitudes observées peuvent ne pas être statistiquement significatives.
En conclusion, on observe que le groupe des femmes ayant des activités bisexuelles est relativement similaire à celui des femmes hétérosexuelles.
Groupe : Hommes bisexuels
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Plaisir sexuel Pénis |
8,86 |
,435 |
63 |
7 |
9 |
|
Plaisir sexuel Prostate |
7,56 |
2,117 |
36 |
0 |
9 |
|
Plaisir sexuel Testicules |
7,48 |
1,649 |
61 |
2 |
9 |
|
Plaisir sexuel Anus |
6,95 |
2,486 |
61 |
0 |
9 |
|
Plaisir sexuel Fesse |
6,49 |
1,523 |
61 |
2 |
9 |
|
Plaisir sexuel Langue |
5,95 |
1,896 |
58 |
0 |
9 |
|
Plaisir sexuel Lèvre |
5,61 |
1,857 |
54 |
2 |
9 |
|
Plaisir sexuel Poitrine |
5,42 |
2,230 |
59 |
0 |
9 |
|
Plaisir sexuel Dos |
5,39 |
1,847 |
54 |
2 |
9 |
|
Plaisir sexuel Ventre |
4,55 |
2,343 |
56 |
0 |
9 |
|
Plaisir sexuel Tête |
3,98 |
2,462 |
49 |
0 |
9 |
Tableau 12 : Classement de l'érogénéité des zones corporelles des 64 hommes bisexuels
L'analyse des moyennes (test de Friedman : n = 29 ; Khi-deux = 154,79 ; ddl = 10 ; p < 0,001 ) indique qu'il existe une différence statistiquement significative entre les moyennes des différentes zones érogènes.
La comparaison des zones érogènes génitales avec les autres zones érogènes, au seuil de significativité de 0,01, donne les résultats suivants (test de Wilcoxon) :
|
|
Note plaisir sexuel Prostate - Note plaisir sexuel Pénis |
Note plaisir sexuel Testicules - Note plaisir sexuel Pénis |
Note plaisir sexuel Anus - Note plaisir sexuel Pénis |
Note plaisir sexuel Fesse - Note plaisir sexuel Pénis |
Note plaisir sexuel Pénis - Note plaisir sexuel Poitrine |
|
Z |
-3,343(a) |
-5,757(a) |
-5,132(a) |
-6,565(a) |
-6,422(b) |
|
Asymp. Sig. (2-tailed) |
,001 |
,000 |
,000 |
,000 |
,000 |
( NB : présentation des comparaisons des moyennes les plus proches ; les autres comparaisons sont significatives à 0,001. )
On observe donc que le plaisir érotique procuré par la stimulation du pénis est le plaisir érotique le plus intense que ces personnes peuvent éprouver. Chez les hommes ayant des activités érotiques bisexuelles, l'hypothèse intermédiaire Hy2 est donc validée.
L'analyse post-hoc exploratoire (Tukey) n'a pas mis en évidence d'autres résultats significatifs. Néanmoins, l'effectif du groupe étant plus faible que celui des hétérosexuels, les similitudes observées peuvent ne pas être statistiquement significatives.
En conclusion, on observe que le groupe des hommes ayant des activités bisexuelles est relativement similaire à celui des hétérosexuels, avec, comme principale différence, une plus grande intensité érogène de la région anale (dans l'ordre : prostate, anus et fesse).
Groupe : Hommes homosexuels
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Plaisir sexuel Pénis |
8,69 |
,704 |
16 |
7 |
9 |
|
Plaisir sexuel Prostate |
8,25 |
1,165 |
8 |
6 |
9 |
|
Plaisir sexuel Anus |
7,58 |
1,676 |
12 |
3 |
9 |
|
Plaisir sexuel Fesse |
6,08 |
1,382 |
13 |
3 |
9 |
|
Plaisir sexuel Langue |
5,83 |
1,697 |
12 |
3 |
9 |
|
Plaisir sexuel Testicules |
5,80 |
2,305 |
15 |
1 |
8 |
|
Plaisir sexuel Lèvre |
5,23 |
1,739 |
13 |
3 |
8 |
|
Plaisir sexuel Poitrine |
5,00 |
2,683 |
11 |
1 |
9 |
|
Plaisir sexuel Ventre |
4,30 |
1,703 |
10 |
2 |
7 |
|
Plaisir sexuel Dos |
3,73 |
1,618 |
11 |
2 |
6 |
|
Plaisir sexuel Tête |
3,64 |
2,098 |
14 |
0 |
7 |
Tableau 13 : Classement de l'érogénéité des zones corporelles des 17 hommes homosexuels
L'analyse des moyennes (test de Friedman : n = 6 ; Khi-deux = 40,44 ; dll = 10 ; p < 0,001 ) indique qu'il existe une différence statistiquement significative entre les moyennes de rang des différentes zones érogènes.
La comparaison des zones érogènes génitales avec les autres zones érogènes donne les résultats suivants (test de Wilcoxon) :
|
|
Note plaisir sexuel Prostate - Note plaisir sexuel Pénis |
Note plaisir sexuel Testicules - Note plaisir sexuel Pénis |
Note plaisir sexuel Anus - Note plaisir sexuel Pénis |
Note plaisir sexuel Fesse - Note plaisir sexuel Pénis |
Note plaisir sexuel Pénis - Note plaisir sexuel Poitrine |
|
Z |
-1,342(a) |
-3,541(a) |
-2,565(a) |
-3,241(a) |
-2,938(b) |
|
Asymp. Sig. (2-tailed) |
,180 |
,000 |
,010 |
,001 |
,003 |
( NB : présentation des comparaisons des moyennes les plus proches ; les autres comparaisons sont significatives à 0,001. )
On observe donc que le plaisir érotique procuré par la stimulation du pénis tend à être le plaisir érotique le plus intense que ces personnes peuvent éprouver. L'absence de différence significative entre les items "pénis" et "prostate" peut être due au petit nombre de données comparées (uniquement 8). Néanmoins, même s'il s'avérait que chez les personnes ayant des activités homosexuelles la prostate était aussi érogène que le pénis, ce résultat ne réfuterait pas l'hypothèse. D'où, chez les hommes ayant des activités érotiques homosexuelles, l'hypothèse intermédiaire Hy2 est donc validée.
L'analyse post-hoc exploratoire (Tukey) n'a pas mis en évidence d'autres résultats significatifs. Néanmoins, le groupe étant relativement petit et la puissance statistique de l'analyse exploratoire étant plus faible, il est difficile de conclure pour ce groupe.
En conclusion, on observe que le groupe des hommes ayant des activités homosexuelles est relativement similaire à celui des bisexuels.
Conclusion
Au terme des analyses statistiques, on observe dans tous les groupes, et tant chez les hommes que chez les femmes, que le plaisir érotique provoqué par la stimulation des zones érogènes des organes génitaux (pénis/clitoris, vagin) est un des plaisirs érotiques les plus intenses que toutes ces personnes peuvent ressentir. L'hypothèse intermédiaire Hy2 est donc vérifiée.
Des analyses complémentaires, réalisées dans les sections suivantes, permettront d'affiner ces résultats.
a.1 – Existe-t-il une différence entre les hommes et les femmes par rapport aux zones érogènes ?
L'exploration graphique des données, à part les deux dernières variables, semble confirmer globalement cette hypothèse.
|
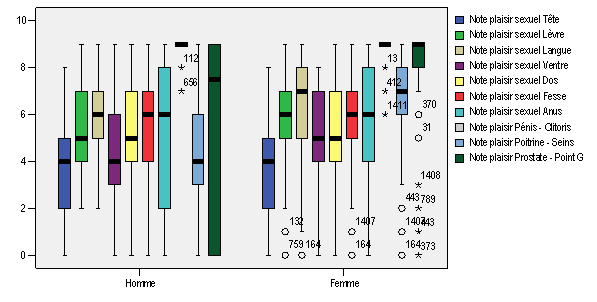
Afin de vérifier si les données des groupes "Homme" et "Femme" sont similaires, les moyennes sont comparées pour chaque variable afin de vérifier s'il existe ou non une différence statistiquement significative entre ces deux groupes.
Les résultats de l'analyse statistique (Mann-Whitney [13] ) sont donnés dans le tableau ci-dessous :
|
|
Note plaisir sexuel Tête |
Note plaisir sexuel Lèvre |
Note plaisir sexuel Langue |
Note plaisir sexuel Ventre |
Note plaisir sexuel Dos |
Note plaisir sexuel Fesse |
Note plaisir sexuel Anus |
Note plaisir Pénis - Clitoris |
Note plaisir Poitrine - Seins |
Note plaisir Prostate - Point G |
|
Mann-Whitney U |
4656,500 |
5970,000 |
5419,500 |
4099,000 |
4446,000 |
5799,000 |
4466,500 |
7629,000 |
2929,000 |
1203,000 |
|
Wilcoxon W |
10327,500 |
11326,000 |
10570,500 |
8104,000 |
8724,000 |
10950,000 |
8561,500 |
14184,000 |
7300,000 |
1944,000 |
|
Z |
-,421 |
-1,152 |
-1,472 |
-1,991 |
-1,193 |
-1,063 |
-,882 |
-,024 |
-6,908 |
-3,168 |
|
Asymp. Sig. (2-tailed) |
,673 |
,249 |
,141 |
,046 |
,233 |
,288 |
,378 |
,981 |
,000 |
,002 |
On observe qu'il n'existe aucune différence statistiquement significative entre les groupes "Homme" et "Femme" pour toutes les régions qui sont anatomiquement similaires : tête, lèvre, langue, dos, fesse et zone anale. La seule exception est le ventre.
Pour les régions sexuellement dimorphes, poitrine-seins, pénis-clitoris, prostate-point G, on observe une différence pour la région de la poitrine, où les seins procurent chez les femmes un plaisir érotique plus intense. A noter que les zones "Prostate" et "Point G" ne sont peut être pas comparables (l'existence du point G est controversée, et de plus les notes données ne correspondent peut être pas toutes à cette zone, dans la mesure où le point G n'est pas forcément connu de toutes les femmes), ce qui pourrait rendre compte de la différence statistique. Quant au vagin et aux testicules, ils ne sont pas comparables ; la nature érogène du vagin pourrait correspondre à la stimulation indirecte du clitoris, qui l'enserre, et les testicules devraient être comparées à leur équivalent embryologique, non inclus dans l'enquête, qui sont les grandes lèvres et partiellement les petites lèvres.
Le résultat le plus intéressant est l'équivalence de l'intensité des sensations érotiques procurées par le pénis et le clitoris, structures ayant la même origine embryologique, et qui sont toutes les deux les zones érogènes à l'origine du plaisir érotique le plus intense.
En conclusion, à part le ventre et surtout les seins, il ne semble pas exister de différences entre les hommes et les femmes par rapport à l'intensité du plaisir érotique provoqué par la stimulation des différentes zones érogènes du corps. Au niveau neurophysiologique, il semble exister, indépendamment du genre, une similitude des réactions érotiques à la stimulation du corps.
a.2 – Zones orgasmogènes
Une analyse complémentaire, relative aux régions corporelles permettant de provoquer l'orgasme, a été effectuée afin d'obtenir des précisions supplémentaires.
NB : Afin de limiter la taille de ce document, ne sont donné ci-dessous que les résultats finaux de cette analyse complémentaire.
|
Item |
Groupe (genre + orientation) |
|
|||||
|
H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
N |
||
|
Orgasme par Tête |
|
2,6% |
0,7% |
5,2% |
|
|
7 |
|
Orgasme par Lèvre |
5,3% |
5,1% |
6,9% |
6,3% |
5,9% |
22 |
|
|
Orgasme par Langue |
8,8% |
5,9% |
8,6% |
6,3% |
5,9% |
28 |
|
|
Orgasme par Poitrine (H) |
5,3% |
|
|
7,8% |
|
8 |
|
|
Orgasme par Seins (F) |
|
14,0% |
19% |
|
|
30 |
|
|
Orgasme par Ventre |
1,8% |
3,7% |
3,4% |
4,7% |
|
12 |
|
|
Orgasme par Pénis (H) |
96,0% |
|
|
98,0% |
100,0% |
189 |
|
|
Orgasme par Testicule (H) |
29,0% |
|
|
44,0% |
12,0% |
63 |
|
|
Orgasme par Clitoris (F) |
|
95,0% |
95,0% |
|
|
184 |
|
|
Orgasme par Vagin (F) |
|
71,0% |
72,0% |
|
|
138 |
|
|
Orgasme par Dos |
0,9% |
2,2% |
3,4% |
3,1% |
|
8 |
|
|
Orgasme par Fesses |
11,0% |
4,4% |
5,2% |
13,0% |
5,9% |
30 |
|
|
Orgasme par Anus |
20,0% |
18,0% |
36,0% |
47,0% |
41,0% |
106 |
|
|
Orgasme par Prostate (H) |
16,0% |
|
|
34,0% |
47,0% |
48 |
|
|
Orgasme par Point G (F) |
0,9% |
48,0% |
60,0% |
|
|
101 |
|
Tableau 14 : Zones orgasmogènes
Remarque : les fréquences supérieures à 25% et à 50% sont grisées claires et foncées, afin de mettre en évidence le profil des réponses.
On observe, tant chez les femmes que chez les hommes, que le penis/clitoris est la principale région orgasmogène.
Les autres régions orgasmogènes sont proches ou liées au pénis/clitoris : testicule pour les hommes et vagin pour les femmes. La prostate et le point G, ainsi que la région anale sont également à l'origine d'orgasme. On observe également que la région anale est plus orgasmogène chez les bisexuels et les homosexuels que chez les hétérosexuels. En plus des zones mentionnées dans le questionnaire, 10 participants indiquent également comme orgasmogène, dans l'ordre des fréquences, les cuisses, les pieds, le cou, les oreilles, les tétons (hommes) et les paupières.
On observe surtout que les principales régions à l'origine de l'orgasme sont situées au niveau de la région pelvienne.
a.3 – Analyse factorielle exploratoire
Une analyse en composantes principales (ACP) a été effectuée afin de rechercher des facteurs qui permettraient de mieux expliciter les résultats. Mais en raison de la diversité des pratiques sexuelles individuelles, des régions corporelles de certains participants à l'enquête ne sont pas stimulées. Ces personnes n'ont donc pas évalués et notés plusieurs items de la question 16. Pour cette raison et afin de pouvoir réaliser l'ACP, les régions du corps les moins stimulées, "Tête" et "Point G" chez les femmes et "Prostate" chez les hommes, n'ont pas été incluses dans l'analyse.
L'ACP sur le groupe "Femmes" met en évidence trois facteurs de valeur propre supérieure à 1. La majorité des items se projetant fortement sur le premier facteur, nous avons décidé de ne conserver qu'un facteur général. Les items qui ont des saturations faibles, à savoir les items "Clitoris", "Vagin" et "Anus", ont été supprimés de l’analyse. La solution retenue, présentée dans le Tableau 15, est composée de 6 items qui expliquent 42,7 % de la variance totale. La consistance interne de ce facteur est satisfaisante ( ? de Cronbach = 0,72 ) : ces items sont homogènes entre-eux.
|
Item du groupe Femmes |
Facteur principal |
|
Plaisir sexuel Lèvre |
,700 |
|
Plaisir sexuel Langue |
,684 |
|
Plaisir sexuel Seins |
,584 |
|
Plaisir sexuel Ventre |
,681 |
|
Plaisir sexuel Dos |
,667 |
|
Plaisir sexuel Fesse |
,596 |
Tableau 15 : Résultats de l'ACP effectuée sur les réponses à la question n° 16, avec le groupe "Femmes" ( N = 141 ).
L'ACP sur le groupe "Hommes" met en évidence trois facteurs de valeur propre supérieure à 1. La majorité des items se projetant fortement sur le premier facteur, nous avons décidé de ne conserver qu'un facteur général. Les items qui ont des saturations faibles, à savoir les items "Pénis", "Testicules" et "Anus", ont été supprimés de l’analyse. La solution retenue, présentée dans le Tableau 16, est composée de 7 items qui expliquent 46,5 % de la variance totale. La consistance interne de ce facteur est satisfaisante : ces items sont homogènes entre-eux ( ? de Cronbach = 0,80 ).
|
Item du groupe Hommes |
Facteur principal |
|
Plaisir sexuel Tête |
,657 |
|
Plaisir sexuel Lèvre |
,752 |
|
Plaisir sexuel Langue |
,740 |
|
Plaisir sexuel Poitrine |
,743 |
|
Plaisir sexuel Ventre |
,689 |
|
Plaisir sexuel Dos |
,652 |
|
Plaisir sexuel Fesse |
,512 |
Tableau 16 : Résultats de l'ACP effectuée sur les réponses à la question n° 16, avec le groupe "Hommes" ( N = 141 ).
Dans les deux groupes, nous interprétons le facteur principal de la manière suivante : ce facteur correspondrait aux zones corporelles les moins érogènes, qui pourraient être les zones érogènes dites "secondaires". On observe que ces zones sont situées en différentes régions du corps, sauf dans la région pelvienne. A partir des résultats de ces analyses factorielles, il semblerait exister deux catégories distinctes de zones érogènes : les zones érogènes pelviennes et toutes les autres zones érogènes. Il apparaît ainsi une tendance qui se retrouve tant chez les femmes que chez les hommes : les principales zones érogènes de la région pelvienne (pénis/clitoris, vagin, testicule, anus) semblent être d'une nature différente que les autres régions érogènes du corps.
a.4 – Analyses complémentaires
Deux analyses complémentaires, relatives au plaisir érotique de la zone anale, sont effectuées afin d'obtenir des précisions supplémentaires.
NB : Afin de limiter la taille de ce document, ne sont donné ci-dessous que les résultats finaux de ces analyses complémentaires.
Plaisir anal et prostatique
Cette analyse complémentaire a été effectuée afin de savoir si le plaisir érotique procuré par les zones érogènes génitales est bien, chez tous les participants, le plaisir érotique le plus intense. La zone anale des hommes et des femmes, ainsi que la prostate, ont été choisies car elles sont dans cette enquête les zones non génitales les plus érogènes. En regroupant toutes les réponses, tant chez les femmes que chez les hommes, chez qui le plaisir procuré par la stimulation de l'anus ou de la prostate est maximal (notes égales à 8 et 9), on observe, comme les moyennes ne sont pas significativement différentes, que le plaisir anal et le plaisir prostatique procurent un plaisir qui est d'une intensité similaire au plaisir procuré par le pénis/clitoris. Ce résultat suggère, pour cet échantillon, que l'intensité du plaisir procuré par la stimulation de l'anus ou de la prostate n'est jamais supérieure à l'intensité du plaisir provoqué par le pénis/clitoris. Néanmoins, les données de l'enquête ne permettent ni de préciser la nature innée ou acquise de cette grande potentialité érogène de l'anus et de la prostate (ou plus précisément de la zone qui a été identifiée par les participants comme étant la prostate), ni de préciser les caractéristiques anatomo-fonctionnelles qui sous-tendent cette particularité.
Érogénicité de la zone anale
On observe chez les hommes homosexuels et surtout bisexuels que la région anale (fesse, anus et prostate) est significativement plus érogène que cette même région pour les hétérosexuels et des femmes bisexuelles (Prostate 7,56 vs 4,89, et Anus 6,95 vs 5,00). Néanmoins, les données de l'enquête ne permettent pas de préciser la nature innée ou acquise de cette particularité.
a.5 – Conclusion
À l'issue de toutes ces analyses statistiques et complémentaires relatives à l'érogénéité des diverses régions du corps, on observe une similitude du pouvoir excitant des différentes zones érogènes entre les hommes et les femmes, et on remarque que les différences les plus importantes n'apparaissent pas en fonction du genre mais de l'orientation des activités érotiques.
On observe également l'existence de 4 grandes zones érogènes, distinctes les unes des autres. Dans l'ordre :
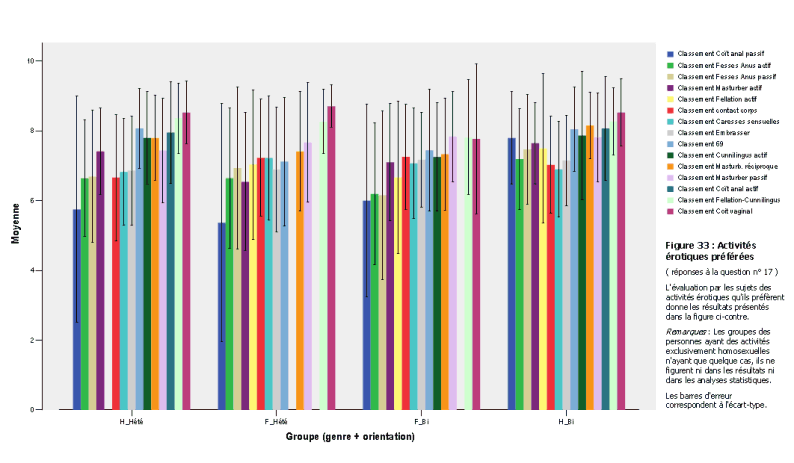
1) La zone génitale, avec principalement le pénis/clitoris et le vagin, et, secondairement, les testicules et le point G ; Cette zone génitale est, dans tous les groupes, significativement distincte des autres zones érogènes, et serait, avec la zone anale, d'une autre nature que les zones non pelviennes.
2) La zone orale, avec principalement la langue, et, secondairement, les lèvres ;
3) la zone anale, avec principalement l'anus, et, secondairement les fesses et la prostate pour les hommes ; L'intensité érogène de cette zone semble dépendre, du moins pour les hommes, de l'orientation des activités érotiques.
4) le tronc (dos et ventre), la tête et les membres.
En conclusion, quel que soit le genre ou l'orientation des activités sexuelles, l'hypothèse intermédiaire Hy2 est toujours vérifiée.
b – Comparaison des activités préférées
( réponses à la question n°17 )
Dans la version finale du questionnaire, l'évaluation par les sujets (87 femmes et 118 hommes) des différentes activités érotiques qu'ils préfèrent donne les résultats suivants (Figure 33) : le coït vaginal et recevoir une fellation ou un cunnilingus sont les activités érotiques préférées pour tous les groupes, sauf les homosexuels. Néanmoins, on observe que les différences sont faibles, en particulier pour les hommes bisexuels, ce qui suggère que le coït vaginal est une activité érotique qui ne se distingue guère des autres.
Les analyses statistiques, réalisées dans les sections suivantes, permettront de vérifier si ces résultats sont significatifs.
Groupe : Hommes hétérosexuels : Activités sexuelles préférées
Les résultats détaillées des réponses du groupe des 77 hommes hétérosexuels sont donnés dans le tableau ci-dessous :
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Classement Coït vaginal |
8,53 |
,895 |
74 |
4 |
9 |
|
Classement Fellation passif |
8,36 |
1,002 |
66 |
4 |
9 |
|
Oro-génital réciproque |
8,07 |
1,150 |
68 |
5 |
9 |
|
Classement Coït anal actif |
7,95 |
1,465 |
41 |
4 |
9 |
|
Classement Cunnilingus actif |
7,80 |
1,324 |
74 |
3 |
9 |
|
Classement Masturb. réciproque |
7,80 |
1,224 |
64 |
4 |
9 |
|
Classement Masturber passif |
7,44 |
1,490 |
71 |
1 |
9 |
|
Classement Masturber actif |
7,41 |
1,234 |
70 |
5 |
9 |
|
Classement Embrasser |
6,87 |
1,563 |
75 |
3 |
9 |
|
Classement Caresses sensuelles |
6,83 |
1,524 |
69 |
4 |
9 |
|
Classement Fesses Anus passif |
6,69 |
1,893 |
42 |
1 |
9 |
|
Classement contact corporel |
6,66 |
1,812 |
71 |
2 |
9 |
|
Classement Fesses Anus actif |
6,64 |
1,669 |
59 |
2 |
9 |
Tableau 17 : Classement des activités érotiques préférées des hommes hétérosexuels
Le résultat du test de Friedman [14] ( n = 20 ; Khi-deux = 55,17 ; ddl = 12 ; p < 0,001 ) indique qu'il existe une différence statistiquement significative entre les moyennes des notes données aux différentes activités érotiques. Les résultats du test de Wilcoxon [15] (cf. tableau ci-dessous), au seuil de significativité de 0,01, indiquent que la préférence pour l'activité du coït vaginal est significativement supérieure aux autres activités, sauf la fellation et le coït anal (cf. cellules grisées).
|
|
Classement Coït vaginal - Classement Fellation passif |
Classement Coït anal- Classement Coït vaginal |
Classement 69 – Classement Coït vaginal |
Classement Coït vaginal - Classement Cunnilingus |
Classement Coït vaginal- Classement Masturber réciproque |
Classement Coït vaginal - Classement Masturber passif |
|
Z |
-,820(a) |
-1,632(b) |
-2,640(b) |
-4,048(a) |
-3,798(a) |
-4,958(a) |
|
Asymp. Sig. (2-tailed) |
,412 |
,103 |
,008 |
,000 |
,000 |
,000 |
( NB : présentation des comparaisons des moyennes les plus proches ; les autres comparaisons sont significatives à 0,001. )
Ces résultats indiquent que les activités érotiques qui provoquent une stimulation intense du pénis (coït anal ou vaginal et fellation) sont équivalentes et sont significativement préférées aux autres activités érotiques.
Groupe : Femmes hétérosexuelles : Activités sexuelles préférées
Les résultats détaillées des réponses du groupe des 56 femmes hétérosexuelles sont donnés dans le tableau ci-dessous :
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Classement Coït vaginal |
8,71 |
,610 |
51 |
6 |
9 |
|
Classement Cunnilingus |
8,26 |
,923 |
53 |
6 |
9 |
|
Classement Masturber passif |
7,67 |
1,717 |
51 |
1 |
9 |
|
Classement Masturb. réciproque |
7,41 |
1,707 |
46 |
1 |
9 |
|
Classement contact corporel |
7,23 |
1,688 |
52 |
2 |
9 |
|
Classement Caresses sensuelles |
7,23 |
1,783 |
53 |
1 |
9 |
|
Oro-génital réciproque |
7,12 |
1,844 |
49 |
1 |
9 |
|
Classement Fellation |
7,04 |
2,139 |
53 |
0 |
9 |
|
Classement Fesses Anus passif |
6,93 |
2,323 |
46 |
0 |
9 |
|
Classement Embrasser |
6,89 |
1,781 |
55 |
2 |
9 |
|
Classement Fesses Anus actif |
6,65 |
2,017 |
37 |
0 |
9 |
|
Classement Masturber actif |
6,54 |
1,982 |
50 |
0 |
9 |
|
Classement Coït anal |
5,37 |
3,419 |
30 |
0 |
9 |
Tableau 18 : Classement des activités érotiques préférées des femmes hétérosexuelles
Le résultat du test de Friedman ( n = 19 ; Khi-deux = 47,77 ; ddl = 12 ; p < 0,001 ) indique qu'il existe une différence statistiquement significative entre les moyennes des notes données aux différentes activités érotiques. Les résultats du test de Wilcoxon (cf. tableau ci-dessous), au seuil de significativité de 0,01, indiquent que les préférences pour les activités du coït vaginal et du cunnilingus sont significativement distinctes entre elles et des autres activités érotiques.
|
|
Classement Coït vaginal - Classement Cunnilingus |
Classement Coït vaginal- Classement Masturber passif |
Classement Masturber passif- Classement Cunnilingus |
Classement Masturber réciproque - Classement Cunnilingus |
Classement 69 – Classement Cunnilingus |
|
Z |
-2,359(a) |
-4,042(a) |
-2,244(b) |
-2,903(b) |
-3,782(b) |
|
Asymp. Sig. (2-tailed) |
,018 |
,000 |
,025 |
,004 |
,000 |
( NB : présentation des comparaisons des moyennes les plus proches ; les autres comparaisons sont significatives à 0,001. )
Ces résultats indiquent que les activités érotiques qui provoquent une stimulation intense du clitoris ou du vagin (coït vaginal, cunnilingus et être masturbé) sont équivalentes et sont significativement préférées aux autres activités érotiques.
Groupe : Femmes bisexuelles : Activités sexuelles préférées
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Classement Masturber passif |
7,83 |
1,289 |
30 |
4 |
9 |
|
Classement Cunnilingus passif |
7,82 |
1,657 |
28 |
3 |
9 |
|
Classement Coït vaginal |
7,77 |
2,156 |
31 |
0 |
9 |
|
Oro-génital réciproque |
7,44 |
1,739 |
27 |
3 |
9 |
|
Classement Masturb. réciproque |
7,33 |
1,605 |
30 |
3 |
9 |
|
Classement contact corporel |
7,26 |
1,505 |
31 |
3 |
9 |
|
Classement Cunnilingus actif |
7,25 |
1,552 |
20 |
4 |
9 |
|
Classement Embrasser |
7,23 |
1,382 |
30 |
5 |
9 |
|
Classement Masturber actif |
7,10 |
1,676 |
29 |
4 |
9 |
|
Classement Caresses sensuelles |
7,07 |
1,585 |
28 |
4 |
9 |
|
Classement Fellation |
6,67 |
2,187 |
30 |
1 |
9 |
|
Classement Fesses Anus actif |
6,19 |
2,040 |
26 |
1 |
9 |
|
Classement Fesses Anus passif |
6,15 |
2,428 |
26 |
0 |
9 |
|
Classement Coït anal |
6,00 |
2,769 |
25 |
0 |
9 |
Tableau 19 : Classement des activités érotiques préférées des 31 femmes bisexuelles
Le résultat du test de Friedman ( n = 13 ; Khi-deux = 29,24 ; ddl = 13 ; p < 0,006 ) indique qu'il existe une différence statistiquement significative entre les moyennes des notes données aux différentes activités érotiques. Les résultats du test de Wilcoxon indiquent que les 4 premières activités du tableau ci-dessus tendent à être préférées aux autres activités érotiques. L'existence de résultats statistiquement non significatifs est peut être dû à la petite taille de l'échantillon. Ces résultats suggèrent que les activités érotiques qui provoquent une stimulation intense du clitoris et du vagin sont préférées aux autres activités érotiques.
Groupe : Hommes bisexuels : Activités sexuelles préférées
|
Item |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Classement Coït vaginal |
8,53 |
,971 |
36 |
5 |
9 |
|
Classement Fellation passif |
8,27 |
,962 |
37 |
6 |
9 |
|
Classement Masturb. réciproque |
8,15 |
,961 |
39 |
6 |
9 |
|
Classement Coït anal actif |
8,07 |
1,486 |
29 |
4 |
9 |
|
Oro-génital réciproque |
8,05 |
1,213 |
39 |
5 |
9 |
|
Classement Cunnilingus |
7,86 |
1,843 |
37 |
0 |
9 |
|
Classement Masturber passif |
7,82 |
1,275 |
39 |
4 |
9 |
|
Classement Coït anal passif |
7,80 |
1,322 |
20 |
5 |
9 |
|
Classement Masturber actif |
7,65 |
1,167 |
40 |
5 |
9 |
|
Classement Fellation actif |
7,50 |
2,147 |
24 |
0 |
9 |
|
Classement Fesses Anus passif |
7,47 |
1,581 |
34 |
3 |
9 |
|
Classement Fesses Anus actif |
7,19 |
1,451 |
36 |
4 |
9 |
|
Classement Embrasser |
7,15 |
1,292 |
40 |
3 |
9 |
|
Classement contact corporel |
7,03 |
1,386 |
39 |
4 |
9 |
|
Classement Caresses sensuelles |
6,90 |
1,374 |
40 |
4 |
9 |
Tableau 20 : Classement des activités érotiques préférées des 41 hommes bisexuels
Le résultat du test de Friedman ( n = 14 ; Khi-deux = 46,69 ; ddl = 14 ; p < 0,001 ) indique qu'il existe une différence statistiquement significative entre les moyennes des notes données aux différentes activités érotiques. Les résultats du test de Wilcoxon indiquent que la préférence pour l'activité du coït vaginal est significativement supérieure aux autres activités, sauf la fellation, l'activité oro-génitale réciproque et le coït anal. Ces résultats indiquent que les activités érotiques qui provoquent une stimulation intense du pénis (coït anal ou vaginal, 69 et fellation) sont équivalentes et sont préférées aux autres activités érotiques.
Groupe : Personnes Homosexuelles: Activités sexuelles préférées
Les groupes des personnes ayant des activités exclusivement homosexuelles n'étant constitués que de quelque cas (4 pour les hommes et 2 pour les femmes), n'ont pas été analysés et ils ne figurent donc ni dans les résultats ni dans les analyses statistiques.
Conclusion
Chez les hétérosexuels, les activités consistant en une pénétration du pénis pour les hommes (coït anal ou vaginal, fellation) et de stimulation du clitoris ou du vagin chez les femmes (coït vaginal et cunnilingus, et secondairement être masturbé) sont significativement préférées aux autres activités érotiques. Mais sur l'ensemble des groupes et surtout pour les bisexuels, on observe peu de différences statistiquement significatives, ce qui suggère que les préférences entre les différentes activités érotiques ne sont pas très importantes.
Néanmoins, il faut noter que les échantillons sont de petites tailles et qu'il existe plus d'une douzaine d'activités à comparer, ce qui affaibli la puissance des tests statistiques. En analysant les tendances des données, il semblerait qu'avec des échantillons de taille plus importante on devrait trouver plus de différences statistiques, montrant une préférence pour les activités qui provoquent une stimulation intense du pénis/clitoris et du vagin (coït vaginal ou anal, recevoir un cunnilingus ou une fellation, être masturbé).
En complément, en analysant les réponses à cette même question, mais données dans les deux premières versions du questionnaire avec un classement sans ex æquo (voir en annexe, page 198), on observe un contraste encore plus fort entre les activités qui provoquent une stimulation du pénis/clitoris et du vagin et les autres activités érotiques.
En conclusion, tous ces résultats suggèrent que les activités érotiques préférées ne sont pas celles qui permettent la reproduction (i.e. les variations autour du coït vaginal), mais celles qui procurent les stimulations les plus intenses aux zones corporelles les plus érogènes. Le coït vaginal, essentiel à la reproduction, est certes une activité préférée, mais parmi plusieurs autres.
c – Le plaisir érotique provoqué par un partenaire est-il supérieur à celui provoqué par autostimulation ?
L'hypothèse intermédiaire Hy3 , "le plaisir érotique provoqué par un partenaire est supérieur à celui provoqué par autostimulation", est testée en utilisant deux questions : une, principale, spécifique à l'intensité du plaisir perçu, l'autre, complémentaire, spécifique aux activités préférées.
( questions n° 18 & 19, en page 9 du questionnaire présenté à l'annexe II )
a – Comparaison des sensations de plaisir érotique en fonction de l'agent inducteur
( réponses à la question n° 18 )
L'évaluation par les sujets de l'intensité du plaisir érotique qu'ils éprouvent, en fonction de l'agent inducteur, indique sans aucune ambiguïté que le plaisir érotique provoqué par un partenaire est supérieur à celui provoqué par autostimulation.
Groupe : Hommes hétérosexuels
|
|
Fréquence |
% |
Cumul % |
|
|
|
Seul |
1 |
,9 |
,9 |
|
Couple |
57 |
50,0 |
50,9 |
|
|
Couple R |
54 |
47,4 |
98,2 |
|
|
Groupe |
1 |
,9 |
99,1 |
|
|
Indifférent |
1 |
,9 |
100,0 |
|
|
Total |
114 |
100,0 |
|
|
Tableau 21 : Agents inducteurs du plaisir le plus intense, chez les 114 hommes hétérosexuels
Par rapport à l'hypothèse à tester, seules 3 modalités de réponses sont importantes :
1) l'autostimulation procure le plaisir érotique le plus intense : 1 cas, soit 0,9% (variable : "Seul") ;
2) la stimulation par une autre personne procure le plaisir érotique le plus intense : 112 cas, soit 98,3% (variables : "Couple" + "Couple Réciproque" + "Groupe") ; et,
3) la situation d'équivalence : 1 cas, soit 0,9% (variable : "Indifférent").
Dans 98,3 % des cas, les sensations provoquées par un partenaire sont plus intenses que celles provoquées par soi-même. Le cas du plaisir par auto-stimulation, qui aurait pu réfuter l'hypothèse, ne représente que 0,9 %.
On observe donc sans aucune ambiguïté que le plaisir érotique provoqué par un partenaire est supérieur à celui provoqué par auto-stimulation. Chez les hommes ayant des activités hétérosexuelles, l'hypothèse intermédiaire Hy3 est validée.
Groupe : Femmes hétérosexuelles
|
|
Fréquence |
% |
Cumul % |
|
|
|
Seul |
5 |
3,7 |
3,7 |
|
Couple |
66 |
48,5 |
52,2 |
|
|
Couple R |
52 |
38,2 |
90,4 |
|
|
Groupe |
1 |
,7 |
91,2 |
|
|
Indifférent |
12 |
8,8 |
100,0 |
|
|
Total |
136 |
100,0 |
|
|
Tableau 22 : Agents inducteurs du plaisir le plus intense, chez les 136 femmes hétérosexuelles
Dans 87,4 % des cas, les sensations provoquées par un partenaire sont plus intenses que celles provoquées par soi-même. Le cas du plaisir par auto-stimulation, qui aurait pu réfuter l'hypothèse, ne représente que 3,7 %.
On observe donc que le plaisir érotique provoqué par un partenaire est supérieur à celui provoqué par auto-stimulation. Chez les femmes ayant des activités hétérosexuelles, l'hypothèse intermédiaire Hy3 est validée.
Groupe : Femmes bisexuelles
|
|
Fréquence |
% |
Cumul % |
|
|
|
Seul |
4 |
6,9 |
6,9 |
|
Couple |
26 |
44,8 |
51,7 |
|
|
Couple R |
18 |
31,0 |
82,8 |
|
|
Groupe |
3 |
5,2 |
87,9 |
|
|
Indifférent |
7 |
12,1 |
100,0 |
|
|
Total |
58 |
100,0 |
|
|
Tableau 23 : Agents inducteurs du plaisir le plus intense, chez les 58 femmes bisexuelles
Dans 81 % des cas, les sensations provoquées par un partenaire sont plus intenses que celles provoquées par soi-même. Le cas du plaisir par auto-stimulation, qui aurait pu réfuter l'hypothèse, ne représente que 6,9 %.
On observe donc que le plaisir érotique provoqué par un partenaire est supérieur à celui provoqué par auto-stimulation. Chez les femmes ayant des activités bisexuelles, l'hypothèse intermédiaire Hy3 est validée.
Groupe : Hommes bisexuels
|
|
Fréquence |
% |
Cumul % |
|
|
|
Seul |
1 |
1,6 |
1,6 |
|
Couple |
26 |
40,6 |
42,2 |
|
|
Couple R |
26 |
40,6 |
82,8 |
|
|
Groupe |
5 |
7,8 |
90,6 |
|
|
Indifférent |
6 |
9,4 |
100,0 |
|
|
Total |
64 |
100,0 |
|
|
Tableau 24 : Agents inducteurs du plaisir le plus intense, chez les 64 hommes bisexuels
Dans 89 % des cas, les sensations provoquées par un partenaire sont plus intenses que celles provoquées par soi-même. Le cas du plaisir par auto-stimulation, qui aurait pu réfuter l'hypothèse, ne représente que 1,6 %.
On observe donc sans aucune ambiguïté que le plaisir érotique provoqué par un partenaire est supérieur à celui provoqué par auto-stimulation. Chez les hommes ayant des activités bisexuelles, l'hypothèse intermédiaire Hy3 est validée.
Groupe : Hommes homosexuels
|
|
Fréquence |
% |
Cumul % |
|
|
|
Seul |
2 |
11,8 |
11,8 |
|
Couple |
5 |
29,4 |
41,2 |
|
|
Couple R |
8 |
47,1 |
88,2 |
|
|
Indifférent |
2 |
11,8 |
100,0 |
|
|
Total |
17 |
100,0 |
|
|
Tableau 25 : Agents inducteurs du plaisir le plus intense, chez les 17 hommes homosexuels
Dans 76,5 % des cas, les sensations provoquées par un partenaire sont plus intenses que celles provoquées par soi-même. Le cas du plaisir par auto-stimulation, qui aurait pu réfuter l'hypothèse, ne représente que 11,8 %.
On observe donc que le plaisir érotique provoqué par un partenaire est supérieur à celui provoqué par auto-stimulation. Chez les hommes ayant des activités homosexuelles, l'hypothèse intermédiaire Hy3 est validée.
Conclusion
Dans tous les groupes, tant chez les hommes que chez les femmes, on observe toujours sans aucune ambiguïté que le plaisir érotique provoqué par un partenaire est supérieur à celui provoqué par auto-stimulation.
L'hypothèse intermédiaire Hy3 est donc validée.
a.1 – Existe-t-il une différence entre les hommes et les femmes par rapport à la préférence de l'agent réalisant l'activité érotique ?
Une hypothèse complémentaire à l'hypothèse principale testée dans cette enquête, est que les réactions érogènes du corps seraient indépendantes du genre et seraient similaires chez les femmes et chez les hommes.
Hétérosexuel Bisexuel
Seul Couple Couple R Groupe Indifférent Seul Couple Couple R Groupe Indifférent
Femme 1 50 48 1 1 7 45 31 5 12
Homme 4 49 38 1 9 2 41 41 8 9
b – Comparaison des activités préférées, en fonction de l'agent
( réponses à la question n° 19 )
L'évaluation par les sujets de l'agent qu'ils préfèrent pour la réalisation de l'activité érotique indique également sans ambiguïté que les personnes préfèrent avoir des activités érotiques avec un partenaire.
Groupe : Hommes hétérosexuels
|
|
Frequency |
Percent |
Valid Percent |
Cumulative Percent |
|
|
|
Seul |
0 |
0,0 |
0,0 |
0,0 |
|
Couple |
113 |
99,1 |
99,1 |
99,1 |
|
|
Groupe |
1 |
,9 |
,9 |
100,0 |
|
|
Total |
114 |
100,0 |
100,0 |
|
|
Tableau 26 : Agents préférés pour les activités érotiques, chez les 114 hommes hétérosexuels
Dans 100 % des cas, les activités érotiques avec un ou plusieurs partenaires sont préférées aux activités érotiques réalisées uniquement par le sujet.
Groupe : Femmes hétérosexuelles
|
|
Frequency |
Percent |
Valid Percent |
Cumulative Percent |
|
|
|
Seul |
1 |
,7 |
,7 |
,7 |
|
Couple |
126 |
92,6 |
92,6 |
93,4 |
|
|
Groupe |
1 |
,7 |
,7 |
94,1 |
|
|
Indifférent |
8 |
5,9 |
5,9 |
100,0 |
|
|
Total |
136 |
100,0 |
100,0 |
|
|
Tableau 27 : Agents préférés pour les activités érotiques, chez les 136 femmes hétérosexuelles
Dans 93,4 % des cas, les activités érotiques avec un ou plusieurs partenaires sont préférées aux activités érotiques réalisées uniquement par le sujet.
Groupe : Femmes bisexuelles
|
|
Frequency |
Percent |
Valid Percent |
Cumulative Percent |
|
|
|
Seul |
2 |
3,4 |
3,4 |
3,4 |
|
|
Couple |
46 |
79,3 |
79,3 |
82,8 |
|
|
Groupe |
2 |
3,4 |
3,4 |
86,2 |
|
|
Indifférent |
8 |
13,8 |
13,8 |
100,0 |
|
|
Total |
58 |
100,0 |
100,0 |
|
Tableau 28 : Agents préférés pour les activités érotiques, chez les 58 femmes bisexuels
Dans 82,7 % des cas, les activités érotiques avec un ou plusieurs partenaires sont préférées aux activités érotiques réalisées uniquement par le sujet.
Groupe : Hommes bisexuels
|
|
Frequency |
Percent |
Valid Percent |
Cumulative Percent |
|
|
|
Seul |
0 |
0,0 |
0,0 |
0,0 |
|
Couple |
46 |
71,9 |
71,9 |
71,9 |
|
|
Groupe |
8 |
12,5 |
12,5 |
84,4 |
|
|
Indifférent |
10 |
15,6 |
15,6 |
100,0 |
|
|
Total |
64 |
100,0 |
100,0 |
|
|
Tableau 29 : Agents préférés pour les activités érotiques, chez les 64 hommes bisexuels
Dans 84,4 % des cas, les activités érotiques avec un ou plusieurs partenaires sont préférées aux activités érotiques réalisées uniquement par le sujet.
Groupe : Hommes homosexuels
|
|
Frequency |
Percent |
Valid Percent |
Cumulative Percent |
|
|
|
Seul |
0 |
0,0 |
0,0 |
0,0 |
|
Couple |
16 |
94,1 |
94,1 |
94,1 |
|
|
Indifférent |
1 |
5,9 |
5,9 |
100,0 |
|
|
Total |
17 |
100,0 |
100,0 |
|
|
Tableau 30 : Agents préférés pour les activités érotiques, chez les 17 hommes homosexuels
Dans 94,1 % des cas, les activités érotiques avec un ou plusieurs partenaires sont préférées aux activités érotiques réalisées uniquement par le sujet.
Conclusion
Dans tous les groupes, tant chez les hommes que chez les femmes, on observe toujours une très nette préférence pour les activités érotiques avec au moins un partenaire.
d – Facteurs à l'origine des activités érotiques
Les résultats présentés dans les chapitres précédants ont montré que le plaisir est un facteur important de l'activité érotique. L'objectif de ce chapitre est d'évaluer l'importance relative du facteur plaisir par rapport à tous les autres facteurs qui interviennent dans l'initiation des activités érotiques avec un partenaire (phase motivationnelle, préalable à la phase consommatoire).
Remarque : comme des analyses préliminaires non présentées dans ce document ont montré que les réponses à cette question sont significativement similaires quelle que soit l'orientation sexuelle, les réponses de toutes les femmes et de tous les hommes ont été regroupées.
a – Résultats
L'évaluation par les sujets de l'importance des différents facteurs à l'origine de leurs activités érotiques avec un partenaire, donne les résultats suivants :
( réponses à la question n° 10, en page 5 du questionnaire présenté à l'annexe II )
Groupe Homme
Les résultats détaillées des réponses du groupe des 113 hommes sont donnés dans le tableau ci-dessous :
|
Item |
Points (Σ notes) |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Raison sexe - Plaisir sexuel |
872 |
8,15 |
1,204 |
107 |
3 |
9 |
|
Raison sexe - Amour |
781 |
8,05 |
1,349 |
97 |
2 |
9 |
|
Raison - Contact physique |
675 |
7,76 |
1,276 |
87 |
4 |
9 |
|
Raison sexe - Complicité |
622 |
7,78 |
1,368 |
80 |
4 |
9 |
|
Raison sexe - Tendresse |
548 |
7,61 |
1,691 |
72 |
2 |
9 |
|
Raison sexe - Pénétrer |
537 |
7,26 |
1,688 |
74 |
3 |
9 |
|
Raison sexe - Pulsion sexuelle |
537 |
7,16 |
1,569 |
75 |
2 |
9 |
|
Raison sexe - Reconnaissance |
462 |
7,57 |
1,617 |
61 |
3 |
9 |
|
Raison sexe - Câlins |
367 |
6,55 |
1,747 |
56 |
2 |
9 |
|
Raison - Demande partenaire |
277 |
6,02 |
2,304 |
46 |
1 |
9 |
|
Raison sexe - Normes sociales |
216 |
6,55 |
2,137 |
33 |
2 |
9 |
|
Raison sexe - Sécurité |
193 |
6,89 |
2,043 |
28 |
2 |
9 |
|
Raison sexe - Chahut |
183 |
6,31 |
2,156 |
29 |
2 |
9 |
|
Raison sexe - Curiosité |
106 |
5,89 |
2,423 |
18 |
2 |
9 |
|
Raison sexe - Enfants |
58 |
4,83 |
2,480 |
12 |
1 |
9 |
Tableau 31 : Facteurs à l'origine des activités érotiques des hommes
On observe que les principaux facteurs donnés par les hommes comme étant à l'origine de leurs activités sexuelles sont, dans l'ordre, le plaisir, l'amour, le contact physique, la complicité, la tendresse, le désir de pénétrer la (ou le) partenaire, une pulsion, le désir d'être reconnu par le partenaire et les caresses sensuelles.
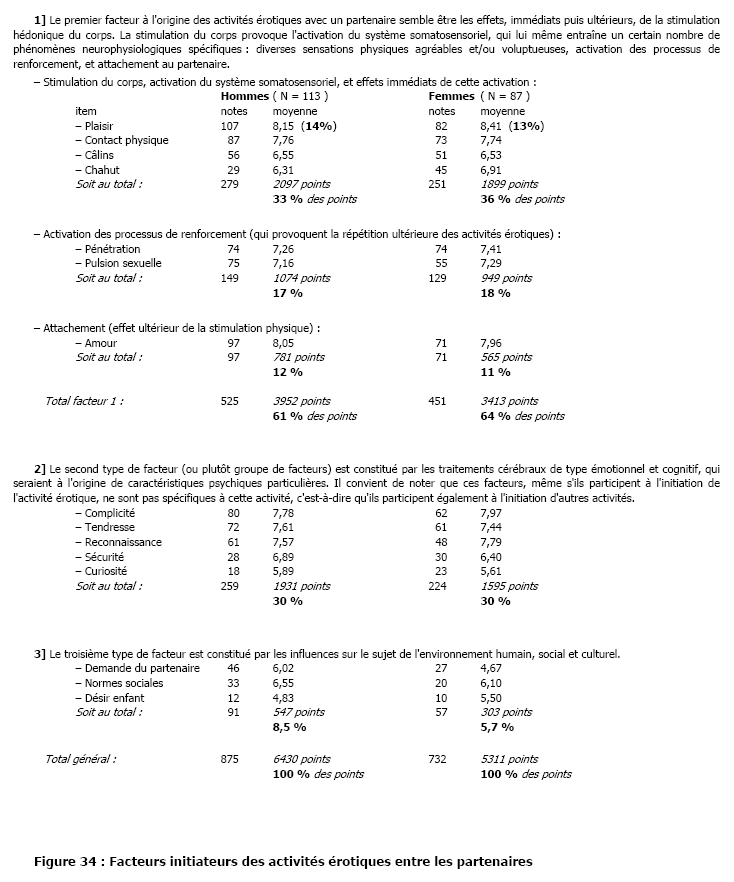
Groupe Femme
Les résultats détaillées des réponses du groupe des 87 femmes sont donnés dans le tableau ci-dessous :
|
Item |
Points (Σ notes) |
Note moyenne |
Écart type |
N |
Minimum |
Maximum |
|
Raison sexe - Plaisir sexuel |
690 |
8,41 |
,831 |
82 |
6 |
9 |
|
Raison sexe - Amour |
565 |
7,96 |
1,870 |
71 |
0 |
9 |
|
Raison - Contact physique |
565 |
7,74 |
1,616 |
73 |
2 |
9 |
|
Raison sexe - Pénétrer |
548 |
7,41 |
1,835 |
74 |
2 |
9 |
|
Raison sexe - Complicité |
494 |
7,97 |
1,402 |
62 |
4 |
9 |
|
Raison sexe - Tendresse |
454 |
7,44 |
1,432 |
61 |
2 |
9 |
|
Raison sexe - Pulsion sexuelle |
401 |
7,29 |
1,641 |
55 |
2 |
9 |
|
Raison sexe - Câlins |
333 |
6,53 |
2,101 |
51 |
2 |
9 |
|
Raison sexe - Reconnaissance |
326 |
6,79 |
2,133 |
48 |
2 |
9 |
|
Raison sexe - Chahut |
311 |
6,91 |
1,905 |
45 |
2 |
9 |
|
Raison sexe - Sécurité |
192 |
6,40 |
2,207 |
30 |
2 |
9 |
|
Raison sexe - Curiosité |
129 |
5,61 |
2,692 |
23 |
1 |
9 |
|
Raison - Demande partenaire |
126 |
4,67 |
2,075 |
27 |
1 |
9 |
|
Raison sexe - Normes sociales |
122 |
6,10 |
3,243 |
20 |
1 |
9 |
|
Raison sexe - Enfants |
55 |
5,50 |
2,635 |
10 |
1 |
9 |
Tableau 32 : Facteurs à l'origine des activités érotiques des femmes
On observe que les principaux facteurs donnés par les femmes comme étant à l'origine de leurs activités sexuelles sont, dans l'ordre, le plaisir, l'amour, le contact physique, le désir d'être pénétrer par le partenaire, la complicité, la tendresse, une pulsion, les caresses sensuelles et le désir d'être reconnu par le partenaire.
On observe également un profil de réponse très similaire entre les femmes et les hommes.
Les réponses brutes des 15 items des raisons des activités érotiques avec un partenaire sont regroupés en fonction du phénomène biologique ou sociologique qui les sous-tend. Les 3 principaux phénomènes retenus sont 1) l'activation du système somatosensoriel, 2) le traitement émotionnel et cognitif, et 3) les facteurs environnementaux externes au sujet.
Pour chaque item sont additionnés le nombre de notes données et les points donnés (points = somme de toutes les notes). Les facteurs principaux à l'origine des activités érotiques avec un partenaire seraient ceux ayant reçu le score le plus élevé (Figure 34).
En conclusion, on observe que les facteurs à l'origine des activités érotiques avec un partenaire sont similaires entre les femmes et les hommes. Le plaisir érotique est un facteur important, mais il n'est pas le seul.
Le facteur principal, d'une importance d'environ 60% (un peu moins de 2/3), serait les effets spécifiques, tant immédiats qu'ultérieurs, de la stimulation du corps. Ce facteur inclut le plaisir érotique (qui représente à lui seul un peu moins de 15%).
Le facteur secondaire, d'une importance d'environ 30% (près de 1/3), serait des caractéristiques émotionnelles et cognitives, non spécifiques à la sexualité.
En schématisant, les deux principaux facteurs qui font que les activités sexuelles avec un (ou plusieurs) partenaires sont nettement préférables aux activités sexuelles avec soi-même, sont : 1) les effets particuliers de la stimulation du corps par autrui, et 2) les effets affectifs de l'interaction positive avec les autres.
Remarque : les réponses à la question des raisons des activités érotiques avec un partenaire peuvent être interprétés de plusieurs manières différentes. L'interprétation présentée ci-dessus est explicitée plus en détail dans la section "discussion", page 105.
b – Existe-t-il une différence entre les hommes et les femmes par rapport aux facteurs initiant l'activité érotique avec un partenaire ?
Une hypothèse complémentaire à l'hypothèse principale testée dans cette enquête, est que les réactions érotiques seraient indépendantes du genre et seraient similaires chez les femmes et chez les hommes.
Somatosensoriel Renforcement Attachement Émotion Environnement
Femme 36 18 11 30 5,7
Homme 33 17 12 30 8,5
Les hommes et les femmes ne semblent donc pas réagir différemment par rapport aux facteurs qui les incitent à avoir des activités érotique avec un partenaire. Au niveau neurophysiologique, il semble exister, indépendamment du genre, une identité des réactions érotiques basiques.
e – Cas analysés à part
Cette section traite de l'analyse des cas qui ont été écartés en raison de l'existence déclarée de troubles physiologiques ou psychologiques qui perturbe la sexualité.
Environ un tiers des participants à l'enquête ont indiqué avoir un problème de ce type. Malheureusement, l'absence de précisions sur le trouble, l'existence de plusieurs troubles simultanés ou la diversité des problèmes indiqués à rendu impossible la constitution de groupes homogènes ayant suffisamment de cas pour permettre des analyses statistiques.
L'analyse descriptive de ces cas indique que 240 personnes (137 femmes et 103 hommes) ont déclaré avoir des troubles qui perturbent actuellement leur sexualité. La prévalence des troubles dans l'échantillon est de 32% au total (240 cas sur 749), de 30% pour les hommes (103 cas sur 348) et de 34% pour les femmes (137 cas sur 398). Ces taux de prévalence sont dans la moyenne de ceux qui sont généralement observés dans les enquêtes concernant les dysfonctions sexuelles : de 10 à 52% pour les hommes et de 25 à 63% pour les femmes (LAUMANN & al. 1999). Les hommes représentent les 2/3 (66%) des 59 personnes ayant un trouble physiologique (39 hommes et 20 femmes), tandis que les femmes représentent également les 2/3 (65%) des 202 personnes ayant un trouble psychologique (117 femmes et 64 hommes). Ces 240 personnes ont déclaré 290 troubles (202 psychiques et 88 physiologiques) et 80 de ces personnes ont déclaré plusieurs troubles. La fréquence des troubles simultanés est de 33% au total (80 personnes sur 240), dont près des 2/3 (59%) chez les femmes (47 femmes et 33 hommes). Les troubles psychologiques représentent plus des 2/3 (70%) des troubles. En résumé, la majorité des hommes ont plutôt des troubles physiologiques et la majorité des femmes ont plutôt des troubles psychiques avec souvent d'autres troubles associés.
Chez les femmes, on observe 38 cas de troubles physiologiques : 15 cas de dyspareunies (douleurs au cours des rapports), 6 cas de vaginisme, 6 cas de sécheresse vaginale, 2 cas d'infections urinaires, 2 cas de traumatisme de la région pelvienne, 2 cas d'obésité et 5 cas divers (problème rénal ou urinaire, accouchement, ...). On observe également 124 cas de troubles psychologiques : 47 cas de gêne importante avec un partenaire, 16 cas d'abus sexuels et 8 cas de viol, 16 cas d'expériences sexuelles négatives (première expérience traumatique, infidélité, inceste entre adulte ...), 14 cas de problèmes d'éducation parentale (négligence, autoritarisme, éducation anti-sexuelle ...), 4 cas de traumatismes dû à des scènes sexuelles choquantes (homosexualité, pornographie ...), 3 cas de violences conjugales, 3 cas de grande gêne provoquée par le questionnaire, 3 cas de pathologie psychique (dépression, anorexie ...), 1 cas de perte de libido, 1 cas de peur des MST, 4 cas divers (avortement, stress ...) et 22 cas non précisés.
Chez les hommes, on observe 50 cas de troubles physiologiques : 22 troubles divers de l'érection, 14 cas d'éjaculation précoce, 5 cas de phimosis, 1 cas de prostatite, 1 cas de trouble de l'éjaculation, 1 cas de douleur pénienne, 1 cas de séropositivité, 2 cas divers (insuffisance rénale ...) et 3 cas non précisés. On observe également 78 cas de troubles psychologiques : 25 cas de gêne importante avec un partenaire, 16 cas d'expériences sexuelles négatives, 8 cas de problèmes d'éducation parentale, 5 cas de pathologie psychique (dépression, schizoïdie ...), 4 cas d'abus sexuel et 2 cas de viol, 2 cas de grande gêne provoquée par le questionnaire, 1 cas de perte de libido, 1 cas de traumatisme dû à des scènes sexuelles choquantes, 1 cas de forte timidité, 1 cas divers et 12 cas non précisés.
On observe en général que les personnes indiquant un problème qui perturbe leur sexualité ont un profil de réponse au questionnaire qui est différent des autres participants. Le plus souvent, l'intensité du plaisir érotique avec un partenaire est inférieure aux autres plaisirs, les personnes préfèrent d'autres activités à l'activité érotique, et, quand elles ont des activités sexuelles, elles préfèrent les sensations et les activités autoérotiques.
|
5 – Discussion
L'objectif de ce travail de recherche était de vérifier si les processus de renforcement, associés aux zones érogènes, étaient bien, chez l'Homme, le facteur primordial à l'origine de l'apprentissage du comportement permettant la reproduction. En utilisant les sensations conscientes de plaisirs comme un indicateur de l'activité des processus de renforcement, la vérification de cette hypothèse à été réalisée au moyen d'un questionnaire destiné à évaluer le rôle et l'importance du plaisir érotique dans le comportement sexuel. Les principaux résultats montrent que 1) la stimulation par un partenaire du pénis/clitoris et du vagin est le plaisir le plus intense que l'être humain peut ressentir ; et 2) les principales activités sexuelles consistent en la stimulation du pénis/clitoris et du vagin. Ces résultats suggèrent que c'est le plaisir érotique (et donc indirectement les processus de renforcement) qui seraient le principal facteur à l'origine des activités sexuelles, dont le coït vaginal qui permet la fécondation.
Le but de ce chapitre est, en tenant compte des différents problèmes rencontrés au cours de cette recherche, d'évaluer la validité des résultats expérimentaux et des hypothèses relatives au comportement de reproduction. La discussion est organisée en 4 parties. Sont successivement discutés : a) les limitations de cette recherche, b) la validité de l'expérimentation, c) les résultats expérimentaux, et d) la validité de l'hypothèse et du modèle.
a – Discussion relative aux limites de cette recherche
Il existe quatre grands types de problèmes généraux qui rendent complexe la vérification, chez l'Homme, de l'hypothèse de l'apprentissage du comportement permettant la reproduction. Ces problèmes sont :
1) L'éthique.
2) Les conditions de vérification non optimales.
3) L'extrapolabilité des données animales à l'Homme.
4) Le manque de connaissances en neurosciences.
Ces différents problèmes sont présentés et discutés dans les paragraphes suivants.
a – L'éthique
L'hypothèse à vérifier est une hypothèse dont la composante fondamentale concerne des processus neurobiologiques de l'organisme humain. Pour vérifier cette hypothèse, il faudrait, d'une part, réaliser différentes expériences complémentaires (lésion focale du noyau accumbens ou de la région préoptique, implantation chronique de canules pour des injections de substances pharmacologiques ...) qui modifieraient de manière irréversible certaines structures cérébrales chez des hommes et des femmes, et, d'autre part, modifier les conditions du développement de plusieurs dizaines de nouveau-nés.
Or on ne peut réaliser des expériences qui portent atteinte à l'intégrité physique ou psychique d'un être humain. Pour cette raison, il est impossible d'effectuer les expériences les plus appropriées, "idéales", qui seraient nécessaires à la vérification expérimentale de l'hypothèse.
La vérification de l'hypothèse est donc faite, par défaut, à partir, d'une part, de données déjà existantes, et, d'autre part, d'expériences humaines éthiquement réalisables, mais moins appropriées et limitées.
Remarque
Pour certains épistémologues, dont Popper (1990), tout ce qui n'est pas vérifiable et réfutable ne relève pas de la science.
Il convient néanmoins de préciser que l'hypothèse de cette étude est une hypothèse scientifique, dans la mesure où elle est vérifiable par le moyen d'expériences. Les expériences appropriées ne sont pas réalisées, non pas parce qu'elles sont irréalisables, mais pour des raisons éthiques, parce que ces expérimentations porteraient atteinte à l'intégrité des éventuels participants.
b – Les conditions de vérification non optimales
Comme les expériences les plus appropriées ne peuvent être réalisées chez l'Homme pour des raisons éthiques, les conditions de la vérification de l'hypothèse ne sont pas optimales.
Les principaux problèmes induits par cette vérification par défaut concernent les limitations des rares vérifications qui peuvent être menées chez l'Homme ainsi que la valeur des données scientifiques déjà existantes.
Problèmes relatifs aux expériences éthiquement réalisables
Pour vérifier l'hypothèse de cette recherche, les expériences réalisables qui ne sont pas contraires à l'éthique sont malheureusement peu nombreuses. Elles correspondent surtout à des observations de situations "naturelles" (telles par exemple la pathologie du spina bifida ou bien des groupes sociaux où la sexualité est taboue) qui s'approchent des conditions expérimentales "idéales".
Ces situations "naturelles" ne correspondent qu'approximativement aux conditions qui seraient requises pour vérifier de manière fiable l'hypothèse de la thèse. De plus, beaucoup de facteurs et de paramètres de ces situations ne sont pas contrôlés, voire sont inconnus. En particulier, il est impossible de connaître et de contrôler la plupart des facteurs qui ont influencé le vécu sexuel des personnes participant à des enquêtes ponctuelles (voir un exemple en annexe IV, page 205).
Pour ces raisons, les données recueillies ne permettent guère de formuler des conclusions valides. Dans le meilleur de cas, il est seulement possible de mettre en évidence des tendances qui peuvent infirmer ou conforter l'hypothèse.
Problèmes relatifs aux données déjà existantes
Pour vérifier l'hypothèse de cette étude, il est également possible d'utiliser des données qui existent déjà et qui proviennent d'expériences relativement proches de l'expérience "idéale". Néanmoins, il existe deux types de problème par rapport à ces données.
Le premier problème est que ces données déjà existantes n'ont pas été recueillies dans le but de vérifier l'hypothèse de cette recherche. Les conditions expérimentales dans lesquelles elles ont été recueillies ne sont pas celles qui auraient été nécessaires pour tester précisément l'hypothèse spécifique de cette étude. De ce fait, ces données sont vraisemblablement différentes de celles qui auraient été recueillies dans l'expérience "idéale". Pour ces raisons, elles ne permettent de tester l'hypothèse que de manière approximative.
Le second problème est qu'une grande majorité de ces données déjà existantes proviennent de l'éthologie ou d'expérimentations animales. Bien qu'un grand nombre de processus neurobiologiques soient commun à tous les mammifères, il est néanmoins souvent difficile de savoir dans quelle mesure une donnée animale est extrapolable à l'Homme.
g – L'extrapolabilité des données animales à l'Homme.
Dans quelle mesure ces données sont-elles extrapolables à l'Homme ?
En effet, les données présentées dans le premier chapitre montrent que la structure du cerveau n'est pas la même des rongeurs à l'Homme, que l'effet des hormones diminue en fonction de la complexité cérébrale, que les phéromones humaines ont un effet faible et essentiellement physiologique, et enfin que le comportement permettant la reproduction chez les rongeurs est très différent de celui des humains.
De plus, les données animales proviennent d'expérimentations en milieu contrôlé, qui, du fait de ce contrôle de certains paramètres, peuvent être des situations bien différentes des situations écologiques normales, voire être appauvries. Ce contexte expérimental particulier influence le développement de l'animal et on observe parfois que l'animal de laboratoire peut être différent structurellement et fonctionnellement de l'animal écologique. Par exemple, l'élevage en isolement des rongeurs supprime le léchage anogénital par la mère qui permet le développement de l'amygdale et du noyau préoptique médian (NPOm) (COOKE & al. 2000 ; voir également Wallen (1996) pour les problèmes induits par le contexte particulier des expériences très connues d'Harlow). D'où le problème de l'extrapolabilité des données animales à l'Homme se complique encore du problème de la validité de ces données animales.
Après analyse, il semblerait même que quasiment aucune donnée animale ne puisse être directement extrapolée à l'Homme.
C'est-à-dire, par exemple, que la mise en évidence de l'importance chez les rongeurs de la testostérone dans la reproduction n'implique absolument pas qu'elle doive également être importante dans les autres espèces de mammifères. En effet, l'expérimentation moléculaire, cellulaire et animale permet surtout de comprendre l'anatomie et le fonctionnement du système nerveux. Elle permet de comprendre que l'effet d'une hormone dépend de l'existence de récepteurs à cette hormone, des modifications neuronales induites par cette hormone et des caractéristiques fonctionnelles de la structure réceptrice. Ainsi, la testostérone ne peut avoir des effets similaires dans une autre espèce que si la structure du système nerveux de cette espèce est similaire à celle des rongeurs. Si la distribution cérébrale des récepteurs change, si les faisceaux de connexion ont une autre architecture, ou si les fonctions des structures neurales réceptrices sont différentes, alors les effets de la même hormone peuvent être radicalement différents.
Or on sait que déjà entre deux espèces très proches, tels les campagnols des prairies et ceux des montagnes, une différence limitée à la distribution cérébrale des récepteurs d'un neuromédiateur peut entraîner d'importantes différences comportementales (INSEL & al. 1992). D'où, connaissant les importantes différences structurelles entre le cerveau des rongeurs, espèce privilégiée de l'expérimentation animale, et celui de l'Homme, il semble difficile, voire impossible, de réaliser des extrapolations directes fiables.
Ce qui semble possible, ce sont des analyses indirectes. L'expérimentation moléculaire, cellulaire et animale permet de comprendre l'anatomie, les fonctions, les rapports structure / fonction et les principes généraux de l'organisation des systèmes neuraux. Ces connaissances permettent, dans un second temps, indirectement, et sous réserve que des connaissances précises relatives à l'organisation structurelle du système nerveux humain soient disponibles, d'expliquer et de prédire certaines caractéristiques fonctionnelles des émotions, de la cognition ou des comportements de l'Homme.
d – Le manque de connaissances en neurosciences
Ainsi, par rapport aux processus de renforcement, même s'il est quasi certain qu'ils sont activés par la stimulation des zones érogènes, on ignore actuellement quels sont précisément les processus exacts qui sont à l'œuvre. D'où certaines expériences cruciales, comme par exemple l'inactivation sélective uniquement des processus de renforcement liés à l'activation sensorielle des zones érogènes, sont aujourd'hui impossibles à réaliser.
Quant aux zones érogènes, on connaît très mal la neurophysiologie érotique du vagin et du point G chez la femme et de la prostate chez l'homme. Il est donc difficile de préciser le rôle de chacune de ces zones dans la dynamique du comportement sexuel.
Ce problème de manque de connaissances, en plus du problème éthique, limite encore le nombre d'expériences possibles, oblige à recourir à des moyens de vérification parfois moins appropriés et plus limités, et rend plus complexe les interprétations des résultats expérimentaux.
Néanmoins, ce problème n'est pas rédhibitoire dans la mesure où il est possible de vérifier l'existence et l'importance des processus de renforcement dans le comportement érotique, tout en ignorant leur localisation précise et le détail des mécanismes neurobiologiques à l'œuvre. L'hypothèse principale de cette recherche ne concerne pas le fonctionnement de ces processus, mais leur intervention dans l'acquisition du comportement permettant la reproduction. La connaissance du fonctionnement précis de ces processus n'est donc pas absolument nécessaire ni à l'élaboration de l'hypothèse, ni à sa vérification. Depuis Olds & Milner, de nombreuses expérimentations ont mis en évidence l'existence de ces processus, qui sont acceptés à la fois par les neurobiologistes et par les psychologues (cf. la théorie de la Récompense / Reward). De plus, il existe des expérimentations qui montrent que ces processus interviennent chez les rongeurs et chez l'Homme dans le comportement de reproduction (voir section "Processus de renforcement", page 46). On dispose donc de données expérimentales qui corroborent l'existence et l'intervention de ces processus dans la sexualité. La vérification de l'hypothèse peut se limiter, dans un premier temps, à tester l'importance – supposée primordiale – des renforcements érotiques par rapport aux autres facteurs connus (hormones sexuelles, phéromones, réflexes sexuels ...). Le fonctionnement détaillé des renforcements érotiques restera spéculatif, mais l'essentiel de l'hypothèse peut être vérifié.
Remarque générale
Par ailleurs, il convient de noter que ce manque de connaissance est en partie dû à des facteurs culturels, et pas uniquement au fait que l'élaboration du savoir scientifique est une activité récente. Il semble en effet, bien que paradoxalement les sociétés occidentales aient une tradition humaniste, que le développement de connaissances objectives relatives à l'Homme ne soit guère une priorité. On observe qu'il existe une recherche "technique" (anatomie, physiologie, psychologie cognitive, pathologie humaine, etc.), mais qu'il n'existe pas de structures institutionnelles ayant pour objet de recherche, par exemple, la compréhension globale de l'être humain, l'étude des différents modes d'organisation sociale ou économique, la réalisation de structures expérimentales d'enseignement ou d'éducation, l'étude de la joie, du bonheur ou encore l'évaluation des valeurs éthiques.
En résumé, il existe apparemment des facteurs psychosociologiques et culturels défavorables aux recherches concernant les aspects les plus essentiels et existentiels de la vie humaine. On observe que tout ce qui concerne l'essence même de la vie humaine n'est pas sujet de recherche. Et même le simple fait de suggérer que la recherche devrait avoir une finalité humaniste semble relever de l'incongruité. Ces caractéristiques psychologiques et culturelle, qui d'ailleurs semblent être une constante des sociétés humaines (EDGERTON 1992), sont un obstacle au développement de connaissances objectives relatives à l'Homme.
e – Remédiations
Les solutions adoptées pour remédier aux problèmes éthiques et de conditions non optimales de vérification sont de réaliser une étude systématique, d'effectuer des observations et des enquêtes, d'utiliser un maximum de données pluridisciplinaires permettant de multiplier les approches et les recoupements, ainsi que de rechercher systématiquement l'absence de réfutation et la maximalisation des corroborations.
Quant au problème de l'extrapolabilité, les solutions adoptées sont d'utiliser principalement des données humaines, de rechercher des observations humaines qui permettent de corroborer ou de réfuter indirectement les données expérimentales animales, de ne pas faire d'extrapolations directes de l'animal à l'Homme, mais de réinterpréter les données animales en fonction de l'évolution phylogénétique, puis d'évaluer leur pertinence par la recherche d'une convergence de similitudes physiologiques, comportementales, ainsi que structurelles et fonctionnelles des systèmes neuraux impliqués.
b – Discussion relative à la validité de l'expérimentation
Quelle est la validité de la partie expérimentale ? Dans quelle mesure les différents problèmes rencontrés affectent-ils les résultats de l'enquête par questionnaire ?
Les principaux problèmes de la vérification expérimentale sont les suivants :
– La dichotomie entre le niveau neurobiologique et le niveau psychologique.
– Les caractéristiques de la population à étudier
– Les critères de représentativité
– La particularité du thème de la sexualité.
a – Niveau biologique & Niveau psychologique
Les données expérimentales spécifiquement recueillies pour vérifier l'hypothèse principale de cette recherche sont collectées à partir d'un questionnaire. Cette méthodologie ne pose pas de problème éthique particulier mais n'est pas la plus appropriée pour tester l'hypothèse.
En effet, le problème est que l'hypothèse principale de ce travail de recherche concerne des processus neurobiologiques (niveau biologique) tandis que les évaluations expérimentales éthiquement réalisables sont basées sur des déclarations conscientes (niveau psychologique). Or actuellement il est très difficile de savoir quels sont les rapports exacts entre les processus neurobiologiques inconscients et les perceptions conscientes de ces processus.
La principale limitation de la vérification des hypothèses, au niveau psychologique, est l'absence de connaissances sur les rapports fonctionnels entre les niveaux biologique et psychologique : il est très difficile de connaître les rapports exacts entre 1] les processus de renforcement activés par la stimulation des zones érogènes (renforcements érotiques), et 2] les sensations conscientes de plaisirs intenses et particuliers (plaisirs érotiques) qui sont perçus lors de la stimulation de ces zones érogènes.
Les principales questions qui découlent de cet absence de connaissances sont :
– Le plaisir érotique correspond-il exactement à l'activation des renforcements érotiques ?
– L'intensité du plaisir érotique correspond-il à l'intensité (ou à la force) du renforcement érotique ?
– Peut-il exister un renforcement érotique sans qu'il y ait perception de plaisir érotique ?
– Inversement, peut-il exister du plaisir érotique en l'absence de renforcement érotique ?
Il semble que l'on peut établir une relation, au moins partielle, entre les renforcements érotiques inconscients et la perception consciente du plaisir érotique. Néanmoins rien ne permet de garantir avec certitude que la sensation consciente de plaisir érotique est un indicateur univoque de l'activité des processus neurobiologiques de renforcement érotique.
D'où les problèmes suivants lors de l'analyse des questionnaires : par exemple lorsqu'une personne déclare n'avoir jamais ressentit du plaisir érotique, cela indique-t-il que ses processus de renforcement érotique n'ont jamais été activés ? Lorsqu'une personne déclare ressentir du plaisir érotique, cela indique-t-il que les processus de renforcement ont été activés ?
Pour répondre à ces questions, on dispose de quelques éléments de réponse :
– Des résultats expérimentaux obtenus principalement chez les rongeurs
– Des résultats d'imagerie cérébrale obtenus chez l'Homme
– Des données psychologiques obtenues par introspection
Les données expérimentales (déjà présentées à la section "Processus de renforcement" du chapitre "e – Apprentissage du comportement permettant la reproduction", p. 45) indiquent que la copulation et l'éjaculation provoquent un renforcement et un état affectif positif chez les mammifères non humains. Chez l'Homme, les données d'imagerie cérébrale indiquent que les régions impliquées dans les renforcements sont activées lors de l'éjaculation et l'orgasme. Ces données suggèrent l'existence d'un phénomène biologique basique et commun à tous les mammifères. Enfin, l'analyse des sensations et des perceptions psychiques humaines, recueillies par introspection, indique que les comportements sexuels, dont surtout le coït et l'éjaculation, procurent du plaisir érotique et sont recherchés pour ce plaisir intense et particulier. Ces données psychologiques suggèrent que les plaisirs érotiques et orgastiques sont fortement impliqués dans les phénomènes de la récompense et des renforcements.
D'où, comme les activités qui provoquent des activations des structures neurales impliquées dans les renforcements provoquent également des sensations conscientes de plaisir érotique, et que ces activités sont ensuite volontairement et activement recherchées pour ce plaisir intense, on peut raisonnablement conclure qu'il existe une forte relation entre l'activité des processus biologiques de renforcement érotique et les sensations psychiques de plaisir érotique. Néanmoins, ces données ne permettent pas de savoir ni si cette relation est directe ou indirecte, ni si elle existe toujours.
En résumé, les problèmes sont les suivants : au niveau psychique, à partir des données présentées juste ci-dessus, on suppose que l'activation des processus de renforcement érotique provoque une sensation consciente de plaisir érotique. D'où, on suppose qu'il pourrait ainsi être possible d'obtenir des informations sur l'activité des processus de renforcement érotique en questionnant une personne sur les différentes sensations de plaisirs érotiques qu'elle perçoit consciemment au cours de sa vie et de ses activités sexuelles.
Néanmoins, cette hypothèse ne repose que sur des recoupements entre des données indirectes provenant de l'expérimentation animale, de l'imagerie cérébrale humaine, des observations comportementales chez l'homme, des perceptions de sensations conscientes et des analyses introspectives. Rien ne permet d'affirmer avec certitude qu'il existe une correspondance univoque entre l'activation des processus de renforcement érotique et les sensations conscientes de plaisir érotique. L'extrapolation d'un certain nombre de conclusion à partir de l'analyse des perceptions d'un sujet de ses sensations de plaisir érotique est donc sujet à caution.
Exprimé autrement, le présupposé, partiellement étayé, sur lequel est bâtit la méthode de vérification est que les sensations de plaisirs érotiques seraient un indicateur de l'activité des processus de renforcement lorsqu'ils sont activés par la stimulation des zones érogènes. Et c'est cet indicateur qui est utilisé pour vérifier, indirectement, l'hypothèse principale. Comme il est actuellement impossible d'évaluer précisément le degré de fiabilité de cet indicateur, il existe donc une marge d'incertitude – que l'on ne peut mesurer – sur la validité intrinsèque de la méthode de vérification.
Quelle est alors la validité de cette méthodologie par questionnaire ?
L'exploration par questionnaire des perceptions conscientes permettra de préciser avec un bon niveau de fiabilité, mais au niveau psychique, le rôle du plaisir érotique dans la dynamique du comportement érotique. Par contre, quelles que soient les conclusions qui pourront être déduites de cette étude, il n'est vraisemblablement pas possible de faire une simple transposition de ces conclusions du niveau psychique au niveau neurobiologique. Il sera donc difficile d'en extrapoler une conclusion absolument fiable concernant le rôle des processus neurobiologiques de renforcement dans l'acquisition du comportement de reproduction.
b – Population à étudier
Le principal – et majeur – problème relatif à la population à étudier est que cette population n'existe plus. En effet, l'objectif de cette recherche est d'identifier les processus primordiaux à l'origine du comportement permettant la reproduction. Or l'influence technologique et culturelle, et surtout la connaissance des activités qui permettent la reproduction, font qu'aujourd'hui il est vraisemblable que les processus neurobiologiques à l'origine du comportementaux de la reproduction sont, au moins partiellement, différents des processus primordiaux. Par exemple, la connaissance de la fonction du coït vaginal fait qu'une personne peut volontairement réaliser cette activité dans un but procréatif : dans ce cas, vraisemblablement, ce sont des processus neurobiologiques télencéphaliques (ganglions de la base, et les cortex frontal, prémoteur et moteur) qui sont principalement à l'œuvre. Au contraire, en situation d'ignorance, ce serait plutôt les processus primordiaux mésencéphaliques et diencéphaliques qui seraient à l'origine d'un comportement involontaire permettant la reproduction.
Pour être certain de limiter l'influence culturelle et d'être dans les conditions les plus proches de celles ayant existé, il faudrait étudier la population des tous premiers homo sapiens, il y a environ 100.000 ans.
Le principal problème de cette étude est que les caractéristiques exactes de cette population originelle sont inconnues et donc spéculatives, et que la population humaine actuelle est certainement très différente pour de nombreux paramètres, dont la plupart sont inconnus.
g – Représentativité de l'échantillon
La représentativité est une caractéristique cruciale de l'échantillon, qui permet à l'issue des analyses d'extrapoler les résultats à l'ensemble de la population étudiée. Il existe plusieurs problèmes spécifiques relatifs aux critères de la représentativité :
– L'existence de différents critères de représentativité.
– L'identification des critères cruciaux et spécifiques à la recherche en cours.
– La répartition et la distribution de ces critères dans la population.
Il existe différents critères de représentativité, variables en fonction des objectifs de la recherche en cours. En effet, il est possible d'avoir des échantillons qui sont représentatifs par rapport aux critères de catégories socio-professionnelles, d'âge et de sexe (ces critères sont les plus utilisés, et sont souvent implicitement requis de manière systématique), mais aussi des croyances religieuses, du régime alimentaire, de la pratique régulière d'activités physiques, de la consommation de substances psychogènes, de la réactivité neurophysiologique, de la densité synaptique du cortex, du système HLA, des groupes sanguins, de certains marqueurs génétiques, etc. L'idéal étant de pouvoir prendre en compte un grand nombre de critères pertinents, de telle sorte qu'on puisse être sûr que l'échantillon est bien, à tout point de vue, représentatif de la population totale. Mais cette situation idéale n'est que rarement possible, et donc, par défaut, on sélectionne l'échantillon en fonction des quelques critères cruciaux qui sont les plus spécifiques et nécessaires pour la recherche en cours, et dont on connaît la distribution dans la population.
Quels sont les critères de représentativité nécessaires et spécifiques à cette recherche ? Contrairement à la majorité des enquêtes concernant les comportements sexuels, l'objectif du questionnaire n'est pas de connaître les pratiques érotiques des personnes interrogées, mais l'importance des sensations de plaisir dans la dynamique du comportement sexuel. Pour donner un exemple précis, une des questions est de savoir quelles sont les zones du corps qui provoquent l'orgasme. Pour cette question, a-t-on besoin du critère de la représentativité professionnelle ? Un ouvrier a-t-il des zones corporelles déclenchant l'orgasme qui sont différentes de celles d'un cadre supérieur ? En fonction des connaissances disponibles, on peut dire que ses opinions politiques seront certainement différentes, l'âge de son premier rapport sexuel sera certainement différent, ses représentations et valeurs sexuelles seront certainement différentes, mais l'orgasme sera-t-il déclenché par le pénis/clitoris chez l'ouvrier et les lèvres et la langue chez le cadre supérieur ? En fonction des connaissances actuelles en sexologie et en neurobiologie, vraisemblablement non. Les critères de représentativité qui sembleraient être les plus pertinents se situeraient au niveau neurophysiologique et seraient tous les facteurs qui influencent d'une manière importante les réactions neurobiologiques érotiques primaires : c'est-à-dire probablement la sensibilité somes-thésique des zones érogènes, la réactivité neurophysiologique, des caractéristiques quantitatives et qualitatives des nerfs qui relient les zones érogènes au cerveau, les caractéristiques hormonales de la puberté, les caractéristiques des processus cérébraux responsables de l'orgasme, etc.
Enfin, une fois les critères cruciaux identifiés, il est nécessaire d'en connaître la distribution dans la population étudiée afin de pouvoir sélectionner un échantillon qui soit représentatif de cette population. Le problème est qu'à part les données de type démographique et socio-économique (âge, sexe, CSP, salaires, etc.), on ne connaît pas la distribution dans la population générale des autres critères possibles (anatomiques, physiologiques, biologiques, taille, poids, régime alimentaire, niveau d'anxiété, consommation de psychotropes, croyances spirituelles, valeurs culturelles et morales, etc.). D'où il est impossible pour de nombreux critère de pouvoir constituer des échantillons représentatifs d'une population donnée. Quant aux problèmes de cette étude, non seulement on ne sait pas si les critères neurophysiologiques cités dans le paragraphe précédant sont réellement pertinents, mais en plus on ne connaît pas la distribution de ces critères dans la population humaine originelle.
La question est donc de savoir si la disparition de la population originelle qui devrait être étudiée et si l'absence de représentativité de l'échantillon par rapport aux caractéristiques neurophysiologiques des réactions sexuelles font que : 1) soit l'étude n'a aucune valeur, ou 2) soit, malgré ces problèmes, les réponses recueillies permettent quand même de formuler des conclusions suffisamment valides pour faire progresser les connaissances relatives au comportement de reproduction.
En effet, on observe que les caractéristiques essentielles de tous les processus basiques et fondamentaux du système nerveux ne sont pas modifiées par les caractéristiques de l'environnement : sensibilité somatique, nociceptive, réflexes, saccades oculaires, peur, curiosité, etc. existent dans toutes les sociétés humaines quel que soit le contexte culturel ou écologique. Et rien ne permet de penser que les réactions neurophysiologiques érotiques de base dérogent à ce constat. Il est donc vraisemblable que la robustesse et l'importance des processus fondamentaux qui ont permis à l'espèce humaine de se reproduire depuis des milliers d'années fassent qu'ils existent chez toutes les personnes et qu'ils ne soient guère influencés par les vicissitudes de la vie quotidienne. Pour ces raisons, les problèmes de représentativité ne seraient pas majeurs. Sauf cas très particuliers (anhédonie, lésion neurale, traumatismes graves, abstinence sexuelle, etc.) qui sont exclus de l'analyse, un échantillon de quelques centaines de personnes des deux sexes et relativement jeunes (18 à 40 ans – âge probable de la procréation à l'époque des origines) devrait permettre d'apporter des éléments de réponse, au niveau psychique, quant au rôle et à l'importance du plaisir érotique dans la dynamique du comportement permettant la reproduction.
d – Particularité du thème de la sexualité.
L'étude du comportement de reproduction humain implique nécessairement l'étude du comportement sexuel. Mais, dans la société occidentale, la sexualité est un sujet très particulier et en partie tabou. Cette particularité est à l'origine de nombreux problèmes, dont certains sont un obstacle à l'étude objective de la sexualité et de la reproduction.
Problèmes généraux de la recherche en sexologie
De manière générale, il n'existe quasiment pas de structures institutionnelles dont l'objectif est l'étude de la sexualité humaine. Les études disponibles sont généralement ponctuelles, limitées à une problématique précise (exemple du SIDA), relèvent d'une approche de type psychosociologique, et sont souvent influencées par des valeurs morales particulières. Les études neurobiologiques restent limitées essentiellement aux rongeurs et concernent la reproduction. Dans ces conditions, comment obtenir des connaissances objectives relatives à l'ensemble des différents phénomènes regroupés dans le concept culturel de "sexualité" ?
On observe également que les recherches relatives à la sexualité sont réalisées le plus souvent à l'initiative d'un homme et non d'une communauté scientifique ou d'un organisme institutionnel. De plus, le thème de la sexualité – ou plutôt un aspect de la sexualité – n'est souvent pour ce chercheur qu'un sujet de recherche parmi d'autres : c'est rarement un spécialiste qui consacre l'essentiel de son activité professionnelle à étudier la sexualité. Cette caractéristique est un obstacle à une recherche approfondie tendant à l'exhaustivité pluridisciplinaire.
Par ailleurs, les éléments et phénomènes regroupés dans la notion culturelle de "sexualité" sont, dans de nombreuses sociétés – dont les sociétés occidentales –, associés à des connotations subjectives et particulières, souvent péjoratives, et sont des sujets facilement polémiques et passionnels. On observe ainsi que la sexualité est un sujet d'étude qui provoque des réactions particulières, tant au niveau social que dans la communauté scientifique. Plus le sujet d'étude est sensible et plus les résultats sont dérangeants, plus les entraves et l'hostilité envers le chercheur augmentent. Cette caractéristique est un sérieux obstacle à la découverte et à la présentation de résultats différents des modèles en usages.
Toujours en raison de la nature particulière de la sexualité, on observe également une influence très forte des valeurs et des représentations culturelles dans la recherche concernant ce sujet. Par rapport aux sujets d'études qui sont conceptualisés et considérés comme "sexuel", ce qui est culturellement valorisé ou stigmatisé, ou bien ce qui est considéré, à une époque donnée, comme étant éthiquement de l'ordre du bien ou mal, influence l'ensemble de la recherche en sexualité. Cette caractéristique est un des obstacles majeurs à l'intention d'objectivation de la recherche scientifique.
On observe aussi que beaucoup d'études relatives à la sexualité concernent ce qui est considéré, à une époque donnée, comme relevant de la pathologie, de la déviance ou de l'immoralité. Il se produit ainsi une sur-représentation d'aspects particuliers au détriment des phénomènes normaux. Cette caractéristique est un obstacle à l'étude et à la compréhension de ce qu'est réellement la sexualité, en dehors de la pathologie et des représentations culturelles négatives.
Enfin, on observe que la recherche relative au comportement de reproduction et à la sexualité est réalisée dans plusieurs champs disciplinaires et sur des sujets d'études a priori difficilement comparables : l'Homme, avec une approche psychologique, sociologique ou ethnologique ; l'animal, en particulier les mammifères inférieurs, avec une approche éthologique et neurobiologique. Cette caractéristique est un obstacle à une compréhension globale de la reproduction et de la sexualité humaine. De plus, ces études relèvent également d'approche le plus souvent monodisciplinaire et analytique, et, de surcroît, concernent généralement des sujets très spécifiques et/ou limités (comportements à risques face aux MST, cooccurrence entre sexualité et consommation de drogue, rôle de la testostérone chez le rat mâle, rôle de l'androsténone (phéromone) dans le déclenchement de la lordose chez la souris femelle, etc.). D'où toutes ces différentes études monodisciplinaires et provenant de différents champs disciplinaires, souvent sans liens entre elles, donnent une vision morcelée et hétéroclite du comportement de reproduction et de la sexualité humaine.
Problèmes particuliers à cette recherche
Par rapport à ce travail de recherche et au recueil d'une partie des données par l'intermédiaire d'un questionnaire, un premier type de problème est la déformation volontaire de la réalité. Déjà, par exemple, on sait que même pour des questions relativement anodines, telle que le nombre de partenaires sexuels au cours de la vie, une partie importante des réponses ne sont pas sincère. D'où, par rapport à des sujets beaucoup plus sensibles, il est quasi certain que plus les activités ou les pensées sexuelles d'une personne s'éloigneront de ce qu'elle considère comme "socialement acceptable", plus la déformation, voire l'autocensure de l'information sera importante. De plus, même si quelques participants à cette étude donneront sur une question sensible des réponses qui sembleront authentiques, rien ne permettra de connaître la proportion exacte de ceux qui ont répondu sincèrement, de ceux qui ont modifié leur réponse, et de ceux qui ont pratiqué l'autocensure. Il est possible que les réponses d'une étude sur la sexualité ne reflètent pas tant les croyances et la pratique sexuelle réelle des personnes, mais plutôt un instantané de ce que le groupe interrogé considère comme "socialement normal et acceptable".
Un second type de problème rencontré dans cette étude, majeur, est l'influence culturelle. Le contexte social et culturel modèle le comportement sexuel, et de ce fait certaines questions ne peuvent être testées de manière fiable. Par exemple, l'orientation "naturelle" de l'activité sexuelle est difficile à évaluer juste à partir de l'observation des comportements sexuels, dans la mesure où en Occident l'importance de l'homophobie et la dominance du modèle hétérosexuel favorisent l'hétérosexualité exclusive.
Un troisième type de problème, également majeur, est que les facteurs qui ont influencé le développement de la sexualité de chaque participant à cette étude ne sont pour la plupart ni connus ni contrôlés. Il est donc très difficile de savoir quels sont les facteurs réellement importants par rapport aux caractéristiques actuelles de la sexualité de ces personnes.
Néanmoins, du fait de l'anonymat et du caractère facultatif de cette enquête par Internet, ainsi que du caractère relativement non problématique des questions principales (évaluer l'intensité du plaisir érotique) on peut supposer que seules les personnes intéressées et peu gênées d'évoquer leur sexualité ont répondu à ce questionnaire de manière honnête et fiable. Cette sur-représentation d'une catégorie particulière de la population ne semble pas être un biais, dans la mesure où ces traits de caractères n'influencent pas la neurophysiologie érotique. La particularité du thème de la sexualité ne semble donc pas trop affecter la validité de cette enquête par questionnaire.
z –Conclusion
Malgré les problèmes méthodologiques exposés dans ce chapitre, la forte relation existant entre le niveau biologique et le niveau psychologique, la robustesse vraisemblable des processus de renforcement érotiques, ainsi que la nature anonyme et facultative du questionnaire, font que les réponses recueillies semblent suffisamment valides pour être exploitées. De plus, l'absence d'ambiguïté des réponses et leur grande cohérence avec l'organisation anatomo-fonctionnelle du système nerveux humain, suggèrent que les hypothèses testées sont apparemment proches de phénomènes neurobiologiques réellement existants. Toutes ces données permettent de conclure que cette enquête et les principaux résultats obtenus ont une validité suffisante pour apporter des éléments de réponses relatifs au comportement de reproduction humain.
c – Discussion relative aux résultats expérimentaux
L'expérimentation, réalisée au moyen d'une enquête par questionnaire, avait pour objectif de vérifier si le plaisir érotique était bien, chez l'Homme, le principal facteur psychique à l'origine du comportement permettant la reproduction (hypothèse Hy ). Afin de tester cette hypothèse, trois hypothèses intermédiaires ont été élaborées de manière à pouvoir réfuter l'hypothèse Hy .
Exprimé autrement, si le plaisir érotique est le facteur majeur à l'origine des activités érotiques, on devait absolument trouver que :
– Hy1 : Le plaisir érotique doit être un des plaisirs les plus intenses que l'organisme peut éprouver.
– Hy2 : Les zones érogènes des organes génitaux (pénis/clitoris, vagin) doivent être à l'origine d'un des plaisirs érotiques les plus intense.
– Hy3 : Le plaisir érotique provoqué par un partenaire doit être supérieur à celui provoqué par autostimulation.
Si ces 3 conditions sont vérifiées, le plaisir érotique (et donc indirectement les processus de renforcement) serait un des facteurs psychiques majeurs à l'origine du développement et de la dynamique du comportement érotique, ainsi que de l'acquisition du coït vaginal reproducteur.
Après la passation du questionnaire, à l'issue de toutes les analyses statistiques, les principaux résultats obtenus sont les suivants : on observe apparemment 4 groupes d'intensité de plaisirs, distincts les uns des autres. Dans l'ordre : 1) le plaisir érotique procuré par un partenaire, qui serait de nature différente aux autres plaisirs naturels ; 2) le plaisir procuré par des émotions ; 3) le plaisir procuré par des processus non émotionnels : les plaisirs auditif, visuel, olfactif, gustatif, intellectuel, corporel et masturbatoire ; et 4) le plaisir procuré artificiellement (par drogues, médicaments ...). Par rapport au plaisir érotique, on observe apparemment 4 grandes zones érogènes, distinctes les unes des autres. Dans l'ordre : 1) La zone génitale, avec principalement le pénis/clitoris et le vagin, et, secondairement, les testicules et le point G ; 2) La zone orale, avec principalement la langue, et, secondairement, les lèvres ; 3) la zone anale, avec principalement l'anus, et, secondairement les fesses et la prostate pour les hommes ; et 4) le tronc (dos et ventre), la tête et les membres. Enfin, dans tous les groupes, tant chez les hommes que chez les femmes, on observe toujours sans aucune ambiguïté que le plaisir érotique provoqué par un partenaire est supérieur à celui provoqué par auto-stimulation.
Par rapport aux activités érotiques, on observe une forte relation entre le plaisir et l'activité : les activités érotiques s'organisent surtout autour des zones les plus érogènes. Les principales activités pratiquées ne sont pas uniquement des variations autour du coït vaginal – bien qu'il s'agisse d'une activité cruciale absolument nécessaire à la reproduction – mais toutes les activités qui stimulent intensément le pénis/clitoris et le vagin. Le plaisir et les renforcements érotiques sont les principaux facteurs qui motivent les activités sexuelles, mais des facteurs affectifs et cognitifs liés aux interactions positives avec le partenaire interviennent également. En conclusion, tous ces résultats suggèrent, tant chez les femmes que chez les hommes, que les activités érotiques qui sont recherchées avec un partenaire ne sont pas celles qui permettent la reproduction (i.e. le coït vaginal), mais celles qui procurent les stimulations les plus intenses aux zones corporelles les plus érogènes. Le coït vaginal, essentiel à la reproduction et à la survie de l'espèce, est une activité préférée, mais parmi plusieurs autres.
Ces principaux résultats, qui valident les hypothèses Hy1 , Hy2 , Hy3 et donc Hy , sont discutés plus en détail dans les sections suivantes afin d'évaluer la portée de leur validité et leur extrapolabilité à la population générale.
a – Le plaisir érotique est-il un des plaisirs les plus intenses que l'organisme peut éprouver ?
Au terme de toutes les analyses des réponses aux questions 13, 1 et 4, on a observé apparemment 4 groupes de nature et d'intensité de plaisir, qui semblent distincts les uns des autres. Dans l'ordre :
1) le plaisir érotique procuré par un partenaire (et qui serait de nature différente aux autres plaisirs naturels) ;
2) le plaisir procuré par des émotions ;
3) le plaisir procuré par des processus non émotionnels : les plaisirs auditif, visuel, olfactif, gustatif, intellectuel, corporel et masturbatoire (sans rappel mnésique des souvenirs du plaisir par partenaire) ;
4) le plaisir procuré artificiellement (drogues, médicaments ...).
Par rapport au plaisir procuré par des émotions, il est à noter que dans certains groupes ce plaisir n'est pas statistiquement différent du plaisir érotique par partenaire. Néanmoins, il n'a pas été examiné dans cette enquête dans quelle mesure l'intensité des émotions positives qui sont considérées comme non sexuelles (bien-être, gaieté, joie, bonheur ...) a été influencé par les effets des activités et surtout des intenses affects sexuels.
Quant aux activités, on observe que toutes les personnes, indépendamment de leur genre ou de leur orientation sexuelle, préfèrent sans ambiguïté les activités érotiques avec un partenaire. En comparant les activités avec les plaisirs, on observe que le plaisir érotique avec un partenaire est le plaisir le plus intense (note la plus élevée, cf. § 4.a.a, p. 68) et que l'activité érotique avec un partenaire est l'activité préférée (note la plus élevée, cf. § 4.a.b, p. 79). Ce résultat suggère fortement qu'il existe une relation causale importante entre le plaisir et l'activité, et corrobore l'hypothèse que le plaisir érotique est un facteur important à l'origine des activités érotiques, en particulier avec un partenaire. Par contre, la relation est moins nette entre le plaisir érotique ressenti, provoqué par soi-même (en général la 2e note la plus élevée, cf. § 4.a.a), et la préférence pour l'activité de masturbation (comprise suivant les groupes entre la 2e et la 7e position, cf. § 4.a.b). Une explication plausible serait qu'une grande partie du plaisir provoqué par la masturbation provienne du rappel mnésique des plaisirs ressentis lors des activités érotiques avec des partenaires ( cf. section "4.a.a.1 – Influence des souvenirs dans l'intensité du plaisir sexuel par auto-stimulation", page 74 ). Cette caractéristique expliquerait le moindre intérêt pour les activités auto-érotiques.
L'ensemble de ces analyses et résultats, en plus de confirmer l'importance des émotions chez l'être humain, indiquent que l'hypothèse Hy1 nécessite des précisions complémentaires. Ce serait uniquement le plaisir érotique procuré par un partenaire qui serait le plaisir "naturel" le plus intense que peut ressentir l'organisme humain. Ce résultat, qui n'était que partiellement prévu mais qui valide néanmoins l'essentiel de l'hypothèse intermédiaire Hy1 , est beaucoup plus significatif pour valider l'hypothèse principale du rôle majeur du plaisir érotique dans l'acquisition de la séquence motrice du coït vaginal. En effet, la masturbation, qui ne permet pas la fécondation, est nettement délaissée au profit des activités érotiques avec des partenaires, qui elles sont susceptibles de permettre la reproduction.
b – Les zones érogènes des organes génitaux sont-elles à l'origine du plaisir érotique le plus intense ?
Au terme de toutes les analyses des réponses aux questions 16 et 17, on a observé, dans les différents groupes et tant chez les hommes que chez les femmes, que le plaisir érotique provoqué par la stimulation des zones érogènes des organes génitaux (pénis/clitoris, vagin) est le plaisir érotique le plus intense que les participants à l'enquête peuvent ressentir.
Néanmoins, même si le pénis est en moyenne la zone corporelle la plus érogène pour les 195 hommes sélectionnés pour les analyses statistiques, on observe que d'autres régions du corps peuvent devenir autant érogènes que le pénis : les testicules (dans 23,6 % des cas), la prostate (dans 18,5 % des cas, mais jusqu'à 25 % des cas chez les hommes ayant des activités bisexuelles), l'anus (18 %, mais 29,7 % pour les bisexuels), les fesses (4,6 %), la poitrine (3,1 %), la langue (5,1 %) et les lèvres (2,7 %). Chez les 194 femmes, le clitoris et le vagin sont en moyenne les zones corporelles les plus érogènes, mais, comme chez les hommes, d'autres régions corporelles peuvent devenir autant érogènes : le point G (dans 40,7 % des cas), les seins (17,0 %), l'anus (16,0 %), les fesses (5,2 %), la langue (7,2 %) et les lèvres (5,2 %) peuvent devenir autant érogènes. On observe même quelques cas où ces autres régions du corps sont plus érogènes que le pénis/clitoris et le vagin. Par ailleurs, par rapport au plaisir orgastique, les zones dont la stimulation peut provoquer l'orgasme sont les mêmes que celles précédemment citées, en particulier celles qui sont les plus érogènes : surtout le pénis/clitoris et le vagin, mais également les testicules, la prostate, le point G et la zone anale, c'est à dire essentiellement des zones localisées au niveau du bassin, tant chez les femmes que chez les hommes. Mais on observe aussi que plusieurs autres régions du corps, telles la tête, le dos ou les pieds, peuvent également provoquer l'orgasme.
Ces résultats indiquent que certaines régions du corps, parfois différentes d'une personne à l'autre, peuvent être très érogènes, voire même dans quelques cas produire un plaisir érotique plus intense que celui provoqué par le pénis/clitoris et le vagin. Ces quelques cas, s'ils ne sont pas des exceptions, pourraient réfuter l'hypothèse. En effet, comme on sait que les pratiques sexuelles changent en fonction du contexte culturel, il est possible de supposer que l'intense érogénéité du pénis/clitoris constatée dans cette étude proviendrait en fait de la pratique culturellement valorisée et dominante du coït vaginal dans les sociétés occidentales ; vraisemblablement, dans des contextes culturels différents, en fonction d'autres pratiques érotiques absentes ou valorisées, on pourrait trouver d'autres régions intensément érogènes. Des données ethnologiques et expérimentales, présentées dans les paragraphes suivants, confirment l'importance des stimulations corporelles dans le développement des zones érogènes. Ainsi, il semble envisageable que de nombreuses régions corporelles puissent devenir hautement érogènes si elles sont adéquatement stimulées, en particulier durant la période cruciale du développement de l'organisme.
Or il est capital pour la validité de l'hypothèse que le pénis/clitoris et le vagin soient les régions corporelles les plus érogènes, et cela quelles que soient les normes et les pratiques culturelles. Une étude plus approfondie de ces résultats et des zones érogènes semble donc nécessaire afin d'évaluer à la fois la validité des réponses recueillies, l'impact des stimulations corporelles et des pratiques sexuelles, ainsi que la pertinence de l'hypothèse.
Développement des zones érogènes
Des données ethnologiques, cliniques et expérimentales montrent que les stimulations précoces sont un facteur important pour le développement des zones érogènes.
Par exemple, chez l'animal, on observe que les rates lèchent régulièrement la région anogénitale de leurs petits et que la déprivation expérimentale de ces stimulations entraîne une augmentation de la latence d'éjaculation chez les rats devenus adultes (MOORE 1984). Ces données suggèrent que la stimulation régulière des organes génitaux (et donc des régions érogènes) durant la période du développement faciliterait le déclenchement des réflexes sexuels.
Chez la femme, par rapport aux seins, contrairement à ce qui est observé dans les sociétés occidentales, on remarque dans les tribus de Nouvelle-Guinée que les enfants reçoivent beaucoup de stimulations somatosensorielles, entre autres au niveau de la poitrine, et que, à l'âge adulte, apparemment toutes les mères ressentent un orgasme lorsqu'elles allaitent des enfants (HERDT & STOLLER 1990). Par rapport aux zones érogènes génitales, en comparant les données provenant, d'une part, des sociétés océaniennes où la sexualité est valorisée et pratiquée dès le plus jeune âge, où les organes génitaux font l'objet de soins et de préparations dès la naissance et où les femmes ont des orgasmes aussi rapidement que les hommes (DIAMOND 2004), et, d'autre part, une société africaine, les So, où le plaisir sexuel est occulté, du moins pour les femmes, et où ces dernières n'éprouvent que de la douleur durant les rapports sexuels (ALLGEIER & ALLGEIER 1992, p.12), il apparaît pour les sujets féminins que certains types de stimulations corporelles et génitales (qui restent à déterminer avec précision) semblent indispensables au développement des processus érotiques génitaux. Ces observations montrent que la région a priori la plus érogène du corps féminin, la zone génitale (clitoris/vagin), peut être soit très érogène soit anérogène en fonction uniquement du contexte développemental, ce qui est un argument fort en faveur d'un rôle majeur de l'apprentissage dans l'érogénéité des zones érogènes. On peut, dans le même ordre d’idée, rappeler l’existence dans plusieurs sociétés traditionnelles de rituels de préparations ou d’ablutions génitales, chez les nourrissons garçons et filles, dans le but de sensibiliser la région sexuelle aux stimulations sensorielles (De RACHEWILTZ 1964). Ces données, ainsi que les données cliniques de l'anorgasmie primaire, vraisemblablement provoquée par l'absence de stimulations génitales (ZWANG in BRENOT 2004) et celles d'observations de sexualité précoce (YATES 1990, 2004), confirment l'importance des stimulations somatosensorielles durant la période du développement. Cette hypothèse de l'influence des expériences érotiques infantiles sur le développement des zones érogènes et des processus érotiques a été initialement proposée par Kinsey A.C. en 1951 et Mead M. en 1955.
Ces données montrent qu'il existe une certaine plasticité des processus impliqués dans les réactions neurophysiologiques érotiques, que les zones érogènes primaires ne sont peut être pas limitées aux régions génitales, et que l'échantillon sélectionné pour l'enquête n'est pas représentatif de toutes les potentialités érogènes de l'organisme humain.
Afin de mieux comprendre la neurophysiologie des zones érogènes et de pouvoir mieux évaluer la validité des résultats de l'enquête, les données anatomiques et neurobiologiques actuellement disponibles sont présentées et discutées dans les paragraphes suivants.
Caractéristiques neurobiologiques des zones érogènes
Les données neurobiologiques actuellement disponibles permettent d'envisager 4 types de structures érogènes périphériques impliqués dans les sensations érotiques.
La première structure périphérique serait constituée par l'association fonctionnelle entre la peau poilue et des fibre C non myélinisées qui ont des relations anatomiques privilégiées avec le cortex insulaire. Ce système serait impliqué dans les sensations tactiles agréables, l'attachement et peut être les sensations érotiques (OLAUSSON & al. 2002 ; WESSBERG & al. 2003). Ce système pourrait participer aux sensations érotiques provenant des régions poilues des zones érogènes, comme par exemple le mont de vénus ou les testicules. Néanmoins, l'importance érotique de ce système semble limitée, dans la mesure où les principales zones érogènes sont constituées de peau glabre.
La seconde structure périphérique pourrait éventuellement être constituée par le tissu de type prostatique, qui se trouve dans la prostate masculine et dans les glandes de Skeve féminine (ZAVIACIC 1998). Ce tissu semble constituer une zone érogène primaire, dont la stimulation provoquerait l'orgasme. La raison de l'existence et la fonction de ce tissu érogène, interne à l'organisme et donc difficile d'accès, sont inconnues. Ce tissus prostatique érogène pourrait expliquer les notes élevées données par les femmes et surtout les hommes bisexuels pour ce qu'ils ont identifié comme étant le point G et la prostate. La faible érogénéité indiquée par les hommes hétérosexuels serait due au fait que le massage prostatique n'est guère pratiqué par les hétérosexuels (REMES 2004).
La troisième structure érogène serait éventuellement le tissus spongieux, qui se trouve dans le pénis masculin et qui constitue le fascia d'Halban chez la femme. Ce fascia, homologue du corps spongieux du pénis, pourrait être à l'origine de l'orgasme vaginal (HOANG & al. 1991).
Le quatrième système périphérique serait constitué par le tissu muco-cutané, qui est un tissu de transition entre la peau externe et les muqueuses internes. Cette peau particulière est caractérisée par une moindre épaisseur du derme et les structures neurales sensitives sont plus proches de l'épiderme que dans les autres types de peau (glabre ou poilue). Les régions corporelles constituées de tissus muco-cutané sont le pénis/clitoris, le prépuce, la partie externe de la vulve, la peau périanale, les mamelons, les lèvres et la conjonctive (WINKELMANN 1959). Ce tissu, associé aux processus de renforcements, pourrait être un système fonctionnel majeur impliqué dans le contrôle de comportements liés aux entrée/sorties de matière de l'organisme (voir la section "e – Généralisation du modèle", page 58). Il semblerait que les caractéristiques anatomiques et fonctionnelle de ce système muco-cutané soient à l'origine des principales zones érogènes primaires. Il permettrait d'expliquer les notes élevées données par les participants à l'enquête pour le pénis/clitoris, les seins, les lèvres et l'anus, ainsi que la désignation de ces zones comme étant les principales permettant de déclencher l'orgasme.
Quant au vagin, les facteurs à l'origine de son intense érogénéité ne sont pas connus avec précision. L'érogénéité pourrait provenir soit de l'innervation intrinsèque du vagin, soit du clitoris (qui enserre le conduit vaginal), soit d'autres structures connexes, soit d'une combinaison de ces possibilités. Les structures connexes seraient le sphincter urétral (LENCK & al. 1992), les glandes de Skene (BENATTAR 2005) et le fascia d'Halban. Ces deux dernières structures, et surtout le fascia d'Halban, pourraient correspondre au controversé point G (HINES 2001). Enfin, il est possible que la stimulation vaginale provoque une stimulation indirecte des structures internes du clitoris, qui entourent le vagin (O'CONNELL & al 1998 ; O'CONNELL & DeLANCEY 2005). Dans ce cas, l'érogénéité du vagin proviendrait en fait de la stimulation indirecte du clitoris. Néanmoins, comme il n'existe pas actuellement d'étude ayant comparé et évalué l'importance relative de ces différentes possibilités de l'érogénéité vaginale, il est difficile de conclure. La seule certitude est que le vagin possède des zones, le plus souvent sur sa paroi antérieure, dont la stimulation tactile est intensément érogène et peut conduire à l'orgasme (ALZATE 1987).
Les facteur à l'origine de l'érogénéité de deux autres zones corporelles, la langue et la conjonctive, sont également mal connus. La langue, indiquée dans l'enquête comme très érogène, n'est pas constituée de tissus muco-cutané. Son érogénéité pourrait provenir de l'existence de récepteurs similaires à ceux du tissu muco-cutané (WINKELMANN 1959) ou du fait que la langue est richement innervée et occupe une surface importante dans les cortex somatosensoriels. La conjonctive, bien qu'étant du tissu muco-cutané, n'est généralement ni considérée ni perçue comme une zone érogène. Néanmoins, un participant à l'enquête indique que sa paupière est une zone orgasmogène.
Les données ci-dessus suggèrent que le tissu muco-cutané serait la structure périphérique qui constituerait les principales zones érogènes primaires. D'où, les caractéristiques de ce tissu permettraient-elles de mettre en évidence des propriétés particulières aux zones les plus érogènes ? Contrairement aux hypothèses de certains auteurs, il n'existerait pas dans ce tissu de récepteurs spécifiques du plaisir érotique (tels par exemple les supposés «corpuscules de la volupté», qui seraient des corpuscules de Meissner modifiés) (MALINOVSKY 1996). Chez tous les primates, on trouve dans le pénis/clitoris les mêmes récepteurs sensoriels que dans les autres structures somatiques (COLD & McGRATH 1999). Par contre, la distribution et l'organisation de ces récepteurs diffèrent : chez l'homme, on observe dans le gland du pénis une grande abondance de terminaisons libres et l'absence de disques de Merkel et des autres mécanorécepteurs typiquement présents dans la peau glabre (doigts par exemple) (HALATA & MUNGER 1986), tandis que dans le prépuce on observe beaucoup de corpuscule de Meissner. Chez la femme, la distribution dans le prépuce et dans le gland du clitoris des terminaisons libres et des récepteurs encapsulés est plus homogène (COLD & TAYLOR 1999). Néanmoins, si ces données montrent des différences structurelles entre les différentes zones érogènes muco-cutanées, on ignore actuellement quels types de différences fonctionnelles elles entraînent. Éventuellement, elles pourraient être à l'origine des différences quantitatives et qualitatives dans les sensations érotiques qui sont produites par chaque zone érogène. C'est un des facteurs qui pourrait expliquer la raison pour laquelle les participants à l'enquête indiquent une gradation des sensations érotiques provenant des zones muco-cutanées : pénis/clitoris en premier, puis seins, lèvres, anus, et, bon dernier, la conjonctive.
Quant à l'érogénéité de ces 4 types de structures érogènes périphériques, il semblerait qu'elle dépende de processus centraux. En effet, on observe lors des sections de la moelle épinière l'apparition de nouvelles zones érogènes (transfert érogène), capable de provoquer des para-orgasmes (SOULIER 2001). Ces données cliniques suggèrent que la nature érogène de certaines stimulations corporelles ne dépendrait pas uniquement de l'existence de récepteurs spécialisés ou d'une organisation particulière des récepteurs au niveau des zones érogènes, mais plutôt d'un traitement effectué à un niveau plus central du système nerveux. Les influx nerveux provoquées par la stimulation de ces tissus muco-cutanés seraient principalement véhiculées par le faisceau spino-thalamique (GUYTON 1989), et aboutiraient au niveau cérébral (région préoptique, MALLICK & al. 1994 ; hypothalamus, YANAGIMOTO & al. 1996). Les données cliniques suggèrent que c'est à ce niveau cérébral que ces influx acquerraient leurs caractéristiques sensorielles érotiques (HEATH 1972).
Enfin, pour terminer l'analyse des facteurs à l'origine de l'érogénéité, il convient de rappeler que les organismes ne seraient pas optimisés (cf. JACOB 1977 ; 1981), qu'il existe de manière significative des variations génétiques (REDON & al. 2006) et épigénétiques (STROHMAN 1997), qu'il existe de surcroît des possibilités d'associations structurelles et fonctionnelles entre les différents processus de l'organisme, et que, pour toutes ces raisons, il n'est peut être pas possible de mettre en évidence un unique et parfait système neuroanatomique qui expliquerait simplement l'érogénéité du corps.
Conclusion
L'étude anatomique et embryologique met en évidence une similitude entre les hommes et les femmes : ce sont apparemment les mêmes structures organiques qui peuvent devenir intensément érogènes : pénis/clitoris, prostate/point G, seins/poitrine, anus, langue et lèvres. Parmi ces zones, trois types de tissus, prostatique, spongieux et surtout muco-cutané, associés à des processus limbiques centraux, seraient à l'origine de la nature érogène primaires de ces zones. La sensibilité érotique de ces zones se développerait en grande partie sous l'effet de stimulations physiques au cours de la période du développement. Les facteurs qui pourraient expliquer les variations inter et intra individuelle entre les différentes zones érogènes seraient l'existence de variations génétiques et épigénétiques, et, surtout, des différences structurelles entre les différentes zones érogènes et des différences dans les pratiques sexuelles, en particulier durant l'enfance et l'adolescence.
Dans le contexte culturel des sociétés occidentales, à partir des résultats de cette étude et surtout des travaux de Masters & Johnson (1966), on observe que le pénis/clitoris et le vagin sont nettement les principales zones érogènes, et de très loin les principales zones orgasmogènes. L'hypothèse Hy2 , "les zones érogènes des organes génitaux sont à l'origine d'un des plaisirs érotiques les plus intenses", est donc, dans ce contexte, validée. Dans les sociétés non occidentales, en fonction des rares données ethnologiques disponibles, on observe que les seins peuvent devenir autant érogènes et orgasmogènes que le clitoris. Pour les autres zones érogènes, l'absence de données ne permet pas de conclure.
À partir des résultats de l'enquête et sous réserve d'un contexte développemental spécifique, il semblerait possible d'envisager que les testicules, le tissus prostatique ou spongieux et l'anus, dont 15 à 30% des participants donnent une note maximale, pourraient être autant, voire peut être plus érogène et orgasmogène que le pénis/clitoris. Néanmoins, comme aucune étude physiologique, clinique ou ethnologique actuellement disponible ne confirme cette hypothèse, cette possibilité reste spéculative.
En conclusion, c'est le pénis/clitoris, le vagin et éventuellement les seins qui sont potentiellement à l'origine du plaisir érotique le plus intense que l'organisme peut ressentir. Et même s'il existerait deux ou trois zones supplémentaires (éventuellement zone anale ou orale) qui pourraient être aussi érogènes et orgasmogènes que les zones génitales, l'hypothèse Hy2 , "les zones érogènes des organes génitaux sont à l'origine d'un des plaisirs érotiques les plus intenses", serait et resterait validée, quelque soit le contexte culturel.
Relation plaisir / activité
Quant aux activités érotiques, on observe que la quasi totalité des personnes préfèrent les activités qui provoquent des stimulations du pénis/clitoris et du vagin, indépendamment de leur genre ou de leur orientation sexuelle. En comparant les activités avec les plaisirs, on observe que les zones génitales sont les plus orgasmogènes (cf. § 4.b.a.2, p. 88), que les plaisirs érotiques génitaux sont les plaisirs érotiques les plus intenses (notes les plus élevées, cf. § 4.b.a, p. 80) et que les activités érotiques génitales sont les activités préférées (notes les plus élevées, cf. § 4.b.b, p. 91). Ce résultat suggère fortement qu'il existe une relation causale importante entre le plaisir et l'activité, et corrobore l'hypothèse que le plaisir érotique est un facteur important à l'origine des activités érotiques, en particulier au niveau des organes génitaux.
Remarque importante : bien qu'on observe que les activités érotiques s'organisent autour des structures les plus érogènes, le pénis et le clitoris, ces activités érotiques ne sont pas spécifiques du coït vaginal. Il est très important de noter que ce coït vaginal, absolument nécessaire à la reproduction, n'est pas toujours l'activité significativement préférée. De plus, il n'a pas été évalué dans cette enquête l'influence de la désirabilité sociale dans la préférence pour le coït vaginal : cette activité a peut être été surévaluée par les participants. Néanmoins, par rapport à ce qui est le plus fondamental, que le coït vaginal soit l'activité érotique préférée ou seulement une des activités préférées, il est pratiqué par tous les hétérosexuels et les bisexuels, ce qui permet d'assurer la reproduction et la survie de l'espèce (voir également la section "5.a.z – Facteurs de l'apprentissage du coït vaginal", page 135 ).
g – Le plaisir érotique provoqué par un partenaire est-il supérieur à celui provoqué par autostimulation ?
La supériorité qualitative des activités érotiques avec un partenaire est également confirmée au niveau physiologique par une plus grande libération de prolactine (+400%), vraisemblable indicateur de la satiété et de la satisfaction sexuelle (BRODY & KRUGER 2006).
En complément, les questions 18 et 19 contiennent plusieurs items destinés à préciser l'hypothèse de la prépondérance des renforcements (ou du plaisir) dans le comportement sexuel, en particulier pour savoir si l'activité érotique se développe en fonction de la recherche du renforcement (ou du plaisir) maximal. En effet, on observe dans quasiment toutes les activités humaines une recherche de la maximalisation des plaisirs : alimentation (gastronomie, diversité des préparations culinaires), audition (diversité de musiques et des ambiances sonores), odorat (parfums, senteurs ...), etc. Par rapport à la sexualité, cette recherche de la maximalisation du plaisir pourrait se traduire par la sommation de diverses stimulations érotiques et hédoniques. C'est vraisemblablement ce qu'on observe au cours des orgies, prisées à certaines époques historiques (PARTRIDGE 2002), où, en plus de la diversité des stimuli érotiques (stimulations par plusieurs partenaires, jeux et danses érotiques, etc.), s'additionnent les stimuli auditifs, visuels, olfactifs et gustatifs provenant de l'ambiance particulière et raffinée, où se mêlent à la décoration suggestive mets fins et boissons capiteuses, musique langoureuse et senteurs enivrantes, afin de fournir de nombreuses, diverses et intenses sensations érotiques et hédoniques. Dans le questionnaire, l'objectif est limité à préciser les éventuels effets du nombre de partenaires au cours des activités sexuelles. L'analyse des réponses indique qu'un peu plus de la moitié des personnes préfèrent les sensations érotiques simples provoquées par un partenaire, et l'autre moitié préfèrent les sensations érotiques provoquées par l'addition des stimulations simultanées et réciproques avec un partenaire. Les personnes préférant les sensations érotiques procurées lors d'activités sexuelles en groupe sont très peu nombreuses. Néanmoins, les internautes sont francophones et la plupart proviennent de France, ou secondairement du Canada, de Suisse et de Belgique, pays occidentaux où le contexte culturel est défavorable à la sexualité de groupe. Cette pratique étant marginale, tant au niveau social que dans l'échantillon, il est donc difficile de tester cette hypothèse complémentaire de la recherche du renforcement érotique et hédonique maximal.
Par ailleurs, le fait que la stimulation érotique par un partenaire soit, de très loin, préférée à l'autostimulation, suggère l'existence d'un phénomène neurophysiologique particulier. L'existence de ce phénomène particulier est de plus corroboré par les résultats de l'analyse factorielle (cf. chapitre "4.a.a.3 – Analyse factorielle exploratoire", page 77 ). Plus généralement, on observe que tous les types d'autostimulation provoquent moins d'effets que les stimulations provoquées par un élément externe à l'organisme. Il est supposé, à partir des études effectuées sur les chatouilles, que les effets de l'auto-stimulation somatosensorielle seraient vraisemblablement inhibés au niveau du cervelet (SELDEN 2004). C'est ce processus neurobiologique qui rendrait compte de la plus grande intensité des stimulations érotiques procurées par un partenaire, comparées aux autostimulations. De manière toute spéculative, on peut supposer que si cette particularité neurophysiologique était inverse (c'est-à-dire si l'autostimulation procurait un plaisir bien plus intense que la stimulation avec un partenaire), il n'y aurait vraisemblablement quasiment pas de comportement sexuel avec autrui, il n'y aurait pas de coït reproducteur, et l'espèce humaine se serait sans doute éteinte depuis longtemps. Il semble dans certain cas qu'il suffise d'une modification neurophysiologique minime pour provoquer des modifications psychiques, comportementales et évolutives majeures. Ce phénomène neurophysiologique particulier et contre-intuitif, qui n'est pas spécifique de la sexualité ou de la reproduction, est conforme au modèle de l'évolution de François Jacob (1977, 1981), qui suppose que les organismes actuels sont le résultat du « bricolage de l'évolution ».
Quant aux activités, on observe sans aucune ambiguïté, tant chez les hommes que chez les femmes, que la quasi totalité des sujets ressentent le plaisir érotique le plus intense avec un partenaire et préfèrent les activités érotiques également avec un partenaire. Il existe ainsi une nette relation entre le plaisir ressenti et l'activité pratiquée, ce qui suggère que le plaisir érotique est un des facteurs importants à l'origine de l'activité érotique.
d – Facteurs à l'origine des activités érotiques
Enfin, la dernière partie de l'enquête concerne l'évaluation de l'importance relative du facteur plaisir par rapport à tous les autres facteurs qui interviennent dans l'initiation des activités érotiques avec un partenaire (question n° 10).
L'analyse des réponses semble mettre en évidence une caractéristique extrêmement intéressante des phénomènes biologiques de la sexualité.
Les 15 items de cette question, ainsi que les réponses à ces items, ont été regroupés en fonction de 3 caractéristiques biologiques et sociales, qui sont bien identifiables et qui jouent un rôle important dans la dynamique des comportements : 1) l'activation du système somatosensoriel, 2) le traitement émotionnel et cognitif, et 3) les facteurs environnementaux externes au sujet.
Sachant que la sexualité consiste au niveau le plus basique en la stimulation érotique des zones érogènes du corps, que les primates sont des animaux de contact, que la stimulation du corps produit des effets physiologiques et psychiques majeurs (voir ci-dessous), et que l'innervation sensorielle du corps constitue le système somatosensoriel (WILLIS & COGGESHALL 1991), nous avons élaboré toutes une série de questions spécifiques aux différentes façons de stimuler le corps et aux effets supposés de l'activation du système somatosensoriel. La stimulation érotique du corps (facteur "plaisir" et item "Pour se donner du plaisir" de la question n° 10 ) recueille 14% des notes chez les hommes et 13% des notes pour les femmes. A priori, par rapport à notre hypothèse de l'importance primordiale du plaisir érotique dans la sexualité, le score est faible et l'hypothèse semble réfutée. En continuant l'analyse des réponses, on trouve que l'ensemble des facteurs correspondant à la stimulation du corps (facteurs "Plaisir", "Contact physique", "Câlins" et "Chahut") recueillent environ un tiers des points (36% pour les femmes et 33% pour les hommes). Ce qui signifie que par rapport à ce que perçoivent les sujets, la stimulation du corps représente un tiers des facteurs qui les incitent à avoir des activités sexuelles avec leur partenaire. Ce résultat, nettement plus important, suggère que le plaisir érotique ne serait qu'un aspect d'un continuum de plaisirs somatosensoriels qui culmineraient éventuellement avec l'orgasme. Il faudrait peut être reformuler l'hypothèse Hy et supposer que les plaisirs somatosensoriels, et non juste le plaisir érotique, seraient à l'origine des activités sexuelles.
En continuant cette analyse, on trouve que ce qui correspondrait aux effets des processus de renforcement (facteurs "Pulsion sexuelle" et "Pénétration" des items "Parce que je ressens comme une tendance ou un besoin qui me pousse à faire des activités sexuelles" et "Pour le désir d'être pénétrée (ou de pénétrer) mon (ou ma) partenaire") représentent environ 17-18% des notes. Ces facteurs "Pulsion sexuelle" et "Pénétration" peuvent être interprétés comme étant la traduction psychique soit d'un instinct de la reproduction, soit des effets des processus de renforcement. L'interprétation relative à l'instinct n'est pas retenue en raison des données du chapitre sur l'innéité (cf. "1.d – Le comportement de reproduction est-il inné ?", p. 13), qui montrent que vraisemblablement il n'existe pas d'instinct de la reproduction. Il est supposé que la stimulation corporelle provoque des renforcements (ayant des effets au niveau des structures prémotrices), et, lorsque ces renforcements sont réactivés, ils seraient perçus comme une pulsion psychique particulière (ce serait un phénomène similaire au fumeur qui ressent une impulsion à fumer en voyant une cigarette ou un cendrier).
L'explication que nous proposons est la suivante : les personnes ont répondu aux facteurs "Plaisir", "Contact physique", "Câlins" et "Chahut" parce qu'elles se rappelaient de ces sensations provoquées au cours de leurs dernières stimulations érotiques. Elles ont répondu aux facteurs "Pulsion sexuelle" et "Pénétration" parce qu'elles se rappelaient d'une impulsion perçue juste avant les activités sexuelles. Les réponses des personnes traduiraient à la fois l'activité inconsciente des processus de renforcement et la perception hédonique consciente de ces processus.
Toujours en continuant l'analyse, on trouve que le facteur "Amour" correspond à environ 11-12% des notes. Ce facteur peut également être interprété de plusieurs façon : en particulier comme étant soit un facteur distinct des processus de renforcement, soit comme étant une des conséquence de l'activité de ces processus. Cette dernière interprétation est retenue dans la mesure où il semblerait que ce qui est culturellement appelé "amour" corresponde en fait chez un sujet à une forme de "dépendance" à certaines caractéristiques des personnes "aimées" (REYNAUD 2005).
Ainsi, en considérant que l'activation des processus de renforcement lors de la stimulation du corps provoque des effets immédiats (les différentes sensations de plaisirs), et des effets ultérieurs (des renforcements, éventuellement au niveau des structures motrices ou prémotrices, et des phénomènes d'attachement romantique ou de dépendance), on peut constituer un super facteur qui regroupe les effets immédiats et ultérieurs, conscients et inconscients, de l'activation des processus de renforcement par les stimulations corporelles. On observe alors que ce super facteur représente 64% des notes chez les femmes et 61% des notes chez les hommes, ce qui signifie que les processus de renforcement (et pas juste le plaisir érotique), indépendamment du sexe, seraient à l'origine de près des deux tiers des facteurs qui incitent les personnes à avoir des activités de plaisirs somatosensoriels (incluant le plaisir érotique et l'orgasme).
Avec une analyse similaire, on constitue un deuxième super facteur, regroupant des facteurs émotionnels et cognitifs (facteurs "Tendresse", "Complicité", "Reconnaissance", "Sécurité" et "Curiosité"). On observe alors que ce super facteur représente 30% des notes chez les hommes et chez les femmes, ce qui signifie que les processus émotionnels et cognitifs, indépendamment du sexe, seraient à l'origine de près d'un tiers des facteurs qui incitent les personnes à avoir des activités sexuelles.
Enfin, le dernier super facteur correspondrait aux influences sur le sujet de l'environnement humain, social et culturel (facteurs "Demande du partenaire", "Normes sociales" et "Désir d'enfants"). Le facteur "désir d'enfants" n'est pas ici considéré comme étant un facteur biologique, dans la mesure où il semble que la naissance d'un enfant est la conséquence du coït vaginal et non d'un désir "instinctif" inné. Lorsqu'un désir d'enfants existe, il proviendrait soit d'un vécu positif au contact d'enfants, soit de l'influence du contexte social et culturel (BADINTER 1980). Ce super facteur correspondant à l'influence de l'environnement est presque négligeable, puisqu'il ne représente que 8,5% des notes chez les hommes et 5,7% des notes chez les femmes.
En conclusion, on observe que les facteurs à l'origine des activités érotiques avec un partenaire sont très similaires entre les femmes et les hommes. Le plaisir érotique est un facteur important, mais c'est surtout l'ensemble des effets, immédiats puis ultérieurs, de l'activation des processus de renforcement par la stimulation du corps qui est principalement à l'origine des activités érotiques.
Ces résultats montrent l'importance du système somatosensoriel, et surtout l'importance des effets de l'activité de ce système, dans la sexualité humaine. En raison de l'importance de ce système, nous allons présenter des données complémentaires qui permettent de mieux comprendre le rôle et l'importance apparemment cruciale des effets – à tous les niveaux de l'organisme – de la stimulation du corps. Toutes les observations montrent que les stimulations somesthésiques, qui ont des effets physiologiques, psychiques et comportementaux, semblent être une véritable nécessité développementale et fonctionnelle chez les mammifères (MONTAGU 1984). Le système somatosensoriel est constitué d’un ensemble de sous-systèmes sensoriels corporels (tact, viscéroception, proprioception, kinesthésie, nociception…), essentiels pour la dynamique fonctionnelle des organismes mammaliens. La somesthésie est le premier système à être fonctionnel au cours de la vie embryonnaire. Le développement des différents récepteurs commence dès la 7e semaine de gestation dans la région péri-buccale, il est achevé pour l'ensemble de la surface cutanée et muqueuse à la 20e semaine, selon une progression céphalo-caudale (LECANUET & al. 1993). Les ganglions rachidiens et les faisceaux, gracile et cunéiforme, se développent à partir de la 8e semaine, le noyau cunéiforme à partir de la 9e semaine (WILLIS & COGGESHALL 1991). Le système somatosensoriel devient alors progressivement fonctionnel à partir du 3e mois de gestation. Ce système est constitué de nombreux récepteurs spécialisés (terminaisons libres, disques de Merkel, corpuscule de Meissner, de Krause, de Pacini et de Ruffini…). Ces récepteurs sont localisés dans le derme, superficiel et profond, dans les muscles, les tendons et les viscères. Ils participent à la genèse des sensations tactiles, thermiques, proprioceptives, kinesthésiques et nociceptives, élaborées à partir d'états spécifiques des tissus de l'organisme (MARIEB 1993). Le système somesthésique est le seul système sensoriel ayant des projections, souvent directes, sur l'ensemble du névraxe : moelle épinière, substance réticulée mésencéphalique et diencéphalique, thalamus, hypothalamus, structures limbiques, cervelet et cortex reçoivent des afférences somesthésiques (WILLIS & COGGESHALL 1991). Ces nombreuses projections rendraient compte des effets physiologiques et comportementaux majeurs produits par les stimulations somesthésiques. Chez les primates, la déprivation des stimuli somesthésiques, en particulier durant la prime enfance, provoque ainsi de nombreux troubles psychiques et comportementaux (cf. les expériences classiques d'Harry F. Harlow avec des macaques rhésus privés de contacts physiques durant les six premiers mois postnataux). Chez l'Homme, la déprivation somesthésique, avec déprivation vestibulaire, est vraisemblablement le facteur principal à l'origine du syndrome d'hospitalisme (SPITZ 1945) et du nanisme psychosocial : croissance retardée, développement psychomoteur et intellectuel perturbé, tristesse, inhibition motrice ou agitation, auto-agressivité et balancement compulsif (ROUBERGUE 2003). De plus, les résultats d'une étude comparative entre plusieurs sociétés préindustrielles semble montrer que la déprivation de stimuli somesthésiques à caractère hédonique (privation de plaisirs somatosensoriels) provoquerait, directement et indirectement, des effets comportementaux et sociaux négatifs : probabilité de sévices physiques et d'un faible niveau d'affection envers les enfants, probabilité d'un statut inférieur de la femme, probabilité de guerre, de torture et d'esclavage, probabilité d'activité religieuse avec des divinités plutôt cruelles et agressives (PRESCOTT 1975). À l'opposé, la stimulation régulière du système somatosensoriel produit de nombreux effets positifs, tant physiologiques, psychiques que comportementaux. Par exemple, on observe chez le nourrisson : un gain pondéral de 47%, à quantité calorique ingérée égale ; une augmentation des performances d'orientation et d'activité motrice ; une diminution de la durée d'hospitalisation, dans le cas de pathologies péri-natales. Et chez l'adulte, une meilleure capacité cytotoxique du système immunitaire ; une diminution des hormones du stress (cortisol et noradrénaline) ; une diminution du niveau d'anxiété ; une diminution de l’état dépressif ; une augmentation de la qualité du sommeil ; un meilleur niveau attentionnel et cognitif ; une facilitation de l'attachement interpersonnel … (FIELD 1995 ; MONTAGU 1979 ; LEIDERMAN & al. 1973). Enfin, à un niveau plus général, on observe que les primates sont des animaux de contact, et que la recherche – voire la nécessité – des stimulations somatosensorielles dépasse largement le cadre de ce qui est culturellement appelé "sexualité". Toutes ces données suggèrent que les fonctions et l'importance du système somatosensoriel sont globales, majeures et cruciales dans la dynamique fonctionnelle de l'organisme humain, et que de ce fait, l'importance de ce système dans le plaisir érotique et la sexualité n'est vraisemblablement pas un hasard.
Quelles sont les limitations et la portée des analyses et des résultats de cette question concernant les facteurs motivant les activités sexuelles ? Les 15 réponses possibles à cette question proviennent de la synthèse des réponses de 2 autres questions testées dans les deux premières versions du questionnaire. Ces 15 réponses, sélectionnées parmi plus d'une trentaine, sont celles qui ont suscité le plus de réponses de la part des participants. Elles sont donc relativement représentatives de ce que le sujet perçoit subjectivement comme étant les raisons de ses activités érotiques. À noter donc que les éventuels facteurs inconscients ne sont pas pris en compte. Par ailleurs, la quasi totalité des participants ayant déjà vécu des activités sexuelles, les réponses ne correspondent vraisemblablement pas aux facteurs qui ont initié les toutes premières activités érotiques. De plus, l'influence des phéromones sexuelles (bien que faible, cf. le chapitre "Olfaction" page 17) n'est pas pris en compte. Enfin, comme la forme finale de cette question (n° 10) n'était présente que dans la troisième version du questionnaire, le nombre de participants est donc plus réduit (113 hommes et 87 femmes). Pour toutes ces raisons, et en tenant compte également du caractère partiellement spéculatif des analyses ci-dessus, les résultats sont donnés sous toutes réserves et nécessitent d'être confirmés par des études ultérieures. Néanmoins, on observe que ces résultats sont très cohérents avec ce qui est connu des processus neurobiologique et de la réalité concrète de la sexualité : les activités sexuelles ne se limitent pas juste aux stimulations érotiques des zones érogènes mais comportent généralement d'autres types de stimulations corporelles ; et lors des activités sexuelles, on observe que les interactions émotionnelles et affectives positives entre les partenaires sont également très importantes. Pour ces raisons, ces résultats semblent suffisamment fiables pour permettent de préciser les caractéristiques et l'importance des différents facteurs impliqués dans le comportement sexuel. Par ailleurs, par rapport aux processus somatosensoriels – qui actuellement ne font guère l'objet de beaucoup de recherches – ces résultats suggèrent que l'étude de ces processus et de leurs effets émotionnels et cognitifs devrait permettre une meilleure compréhension de la sexualité et de l'affectivité humaine.
Néanmoins, même sujet à cautions, ces résultats montrent que les renforcements érotiques (ou le plaisir érotique) n'est pas le facteur principal de la phase motivationnelle du comportement sexuel. Ce serait plutôt les processus de renforcement activés par l'ensemble du système somatosensoriel ("renforcements somatosensoriels", ou, au niveau psychique, les "plaisirs somatosensoriels") qui seraient le principal facteur incitant les partenaires à avoir des activités sexuelles. Pour ces raisons, l'hypothèse principale, rappelée ci-dessous, nécessite des précisions complémentaires.
H : Niveau (neuro)biologique : L'acquisition du comportement permettant la reproduction dépend – principalement mais indirectement – de l'activation des processus de renforcement, provoquée par la stimulation des zones érogènes des organes génitaux (Figure 22, page 50).
Hy : Niveau psychologique : L'acquisition du comportement permettant la reproduction dépend – principalement mais indirectement – de la recherche de sensations conscientes de plaisir érotique, provoquées par la stimulation des zones érogènes des organes génitaux. (Figure 26, page 62).
Les précisions complémentaires seraient les suivantes (les modifications sont indiquées en gras) :
H : Niveau (neuro)biologique : L'acquisition du comportement permettant la reproduction dépend – principalement mais indirectement – de l'activation des processus de renforcement, provoquée par la stimulation du corps et des zones érogènes des organes génitaux.
Hy : Niveau psychologique : L'acquisition du comportement permettant la reproduction dépend – principalement mais indirectement – de la recherche de sensations conscientes de plaisirs somatosensoriels, provoquées par la stimulation du corps et des zones érogènes des organes génitaux.
Ces précisions ne remettent pas en cause la caractéristique essentielle de l'hypothèse, qui est le rôle primordial des processus de renforcement dans l'apprentissage à la fois du comportement permettant la reproduction et de la séquence cruciale du coït vaginal. Cependant, ce serait les renforcements somatosensoriels, et non juste les renforcements érotiques, qui seraient primordiaux dans la phase motivationnelle du comportement sexuel. Pour la phase consommatoire, les renforcements érotiques semblent toujours être les seuls facteurs à l'origine de la focalisation des activités érotiques sur les zones érogènes.
Il semblerait donc exister un phénomène biologique particulier qui apparaîtrait lors de la stimulation du système somatosensoriel, et qui serait constitué par tous les effets immédiats, puis ultérieurs, de la stimulation du corps. Ce qui est culturellement appelé "sexualité" ne correspondrait qu'a une partie de ce phénomène.
e – Validité des réponses et des résultats
L'objectif de cette section est : 1) d'évaluer la validité des réponses données par les participants à l'enquête ; et, 2) d'évaluer la validité des résultats de l'analyse de ces réponses.
Après la lecture, l'examen et la saisie informatique des réponses au questionnaire, l'impression générale est que la grande majorité des participants ont répondu de manière très sérieuse et appliquée. Ce fait s'explique vraisemblablement par la longueur du questionnaire, qui a induit une sélection où seules les personnes très intéressées ont répondu. Ce biais ne semble pas être un problème, car il n'affecterait pas les réponses : aucune donnée connue ne permet de dire que l'intérêt pour un questionnaire ou pour la sexualité influence les processus basiques des réactions érotiques. Néanmoins, un petit groupe de participants, peut être moins consciencieux, n'a pas donné de réponses à plusieurs questions, et quelques participants n'ont apparemment pas compris les consignes. Tous ces cas ont été exclus de l'analyse des réponses.
Le problème identifié le plus notable est la fiabilité de la notation. Les réponses aux principales questions se font par l'intermédiaire de notes allant de 0 à 9, et, à partir des quelques cas où il a été possible de réaliser deux ou trois passations successives aux mêmes personnes, il est apparut que seul les 2 ou 3 notes les plus extrêmes (les plus intenses et les moins intenses) sont relativement fiables. Les notes intermédiaires fluctuent à ± 1 ou 2 points. Néanmoins, ce problème n'est pas gênant dans la mesure où la validation ou la réfutation des hypothèses est basée sur les notes les plus élevées et pas sur les notes intermédiaires.
Quant au profil hédonique et comportemental des participants qui est apparu à la lecture des réponses, il ne met en évidence aucun problème particulier. Pour les aspects qui sont comparables, les réponses sont globalement similaires et compatibles avec les données connues en sexologie (ZWANG in BRENOT 2004 ; ALLGEIER & ALLGEIER 1992 ; MASTERS & JOHNSON 1966 ; LAUMANN & al. 1999 ; RÉMÈS 2004). Par exemple, les régions du corps qui ne seraient jamais érogènes, telles le front, les joues, le nez, les oreilles, la face postérieure des épaules, des coudes, la face antérieure des genoux, la plante des pieds, etc. (ZWANG in BRENOT 2004), ne sont quasiment jamais citées par les participants.
En conclusion, l'évaluation de la validité des réponses ne met pas en évidence de problème majeur susceptible de rendre inutilisable les données recueillies au cours de cette enquête.
Quant aux résultats provenant de l'analyse de ces réponses, le principal problème est qu'il a fallu, au moins initialement, distinguer plusieurs groupes (homme, femme, hétérosexuel, bisexuel, homosexuel, anorgasmique, etc.). De plus, l'existence de 3 versions du questionnaire (mais concernant uniquement des questions complémentaires) ainsi que parfois l'absence de réponses à une partie des questions diminuent encore le nombre de cas pouvant être pris en compte dans certaines analyses. Pour ces raisons, le nombre de sujets par groupe est au final relativement faible par rapport au nombre total de participants. Plusieurs groupes n'ont pas pu être analysés statistiquement et seul deux groupes contiennent plus de 100 individus, malgré un échantillon initial de 749 personnes. D'où, pour les petits groupes, la portée des conclusions est plus limitée.
Néanmoins, l'analyse de tous les résultats montre qu'ils sont cohérents entre-eux, et, quand la comparaison est possible, qu'ils sont conformes aux connaissances anatomiques et fonctionnelles du système nerveux, ainsi qu'aux données des sciences humaines concernant la sexualité. En particulier, la conformité aux connaissances sexologiques des deux principaux résultats de l'enquête, à savoir que le plaisir le plus intense que l'organisme peut ressentir provient de la stimulation des organes génitaux par un partenaire et que les facteurs à l'origine des activités érotiques humaines sont principalement le plaisir et des émotions (tendresse, complicité ...), suggère fortement l'absence de biais ou d'erreurs d'analyse.
En conclusion, l'étude des résultats ne met pas en évidence de problème majeur susceptible d'invalider les résultats principaux, qui sont ceux relatifs à l'importance du plaisir érotique (questions 1, 4, 13, 16, 17, 18, 19). Pour une partie des autres résultats provenant de l'analyse de petits groupes, et en particulier pour les résultats apparemment très intéressants relatifs aux facteurs à l'origine du désir d'avoir des activités sexuelles avec un partenaire (question 10), des expérimentations complémentaires seraient souhaitables.
z – Généralisation des résultats
Facteurs de l'apprentissage du coït vaginal
Une question absolument cruciale pour la reproduction est de savoir comment est appris le coït vaginal. Le modèle théorique et les résultats de l'enquête permettent-ils d'apporter des éléments de réponse ?
Actuellement, et sans doute depuis l'époque des origine, le coït est appris par observation, imitation ou par initiation. Mais pour les tous premiers Hommes ? Car en l'absence de processus innés et en l'absence de connaissances innées, la séquence motrice du coït vaginal est a priori improbable : l'orifice vaginal n'est guère visible et sa profondeur n'est nullement apparente, le vagin est moins érogène et surtout moins accessible que le clitoris, la conception intellectuelle de l'intromission n'est pas évidente, et la pénétration du pénis dans le vagin juste au hasard des stimulations érotiques est plus qu'aléatoire. Alors, quels seraient donc les facteurs à l'origine de la découverte et de l'apprentissage de ce coït vaginal ? Même si l'on ignore tout du contexte ayant existé aux origines de l'espèce humaine, la réponse – même théorique – à cette question fondamentale est importante, au minimum pour la capacité explicative du modèle. Si ce modèle peut rendre compte de la découverte cruciale du coït vaginal dans le contexte le plus défavorable, c'est-à-dire sans aides internes (instinct, réflexes sexuels, etc.) ou externes (par exemple l'observation du coït d'autres animaux), mais uniquement à partir des éléments constitutifs du modèle, à savoir les processus de renforcement, les zones érogènes, la morphologie complémentaire des organes génitaux et des caractéristiques émotionnelles et cognitives de l'organisme humain, alors sa crédibilité sera plus grande.
En fonction des données sexologiques (MASTERS & JOHNSON 1980 ; ALLGEIER & ALLGEIER 1992), on observe que le pénis/clitoris (ou plutôt la vulve pour la femme) est à la fois la région la plus érogène ainsi qu'une région visible et facilement accessible. Ces caractéristiques anatomiques et physiologiques seraient à l'origine du développement et de l'organisation, autour de ces zones les plus érogènes, de la majorité des activités érotiques. Mais ces activités génitales ne semblent pas spécifiques du coït vaginal. Ce qui serait recherché n'est pas le coït vaginal, mais, grâce à la diversité des activités morphologiquement possibles, la maximalisation des plaisirs érotiques procurés par la stimulation du pénis/clitoris. C'est ce qu'on observe dans les réponses au questionnaire, où les activités érotiques préférées (coït vaginal, fellation/cunnilingus par la/le partenaire, masturbation par le partenaire) ne sont pas spécifiques du coït vaginal mais de la stimulation du pénis/clitoris, qui sont les zones déclarées comme les plus érogènes.
Néanmoins, on observe chez les Bonobos et chez l'Homme dans les sociétés où il existe peu de restrictions culturelles, que les activités érotiques sont fréquentes (plusieurs fois par jour) et débutent dès les premières années de la vie (HASHIMOTO 1997 ; DE WAAL 1992, 1996, 1990 ; SUGGS 1966 ; MALINOWSKI 1970 ; HENRY & HENRY 1974). D'où, entre l'âge de 4-5 ans et la puberté, vers 15 ans, il existe une période d'une dizaine d'années de pratiques représentant une potentialité de plusieurs milliers d'activités sexuelles. De surcroît, les études psychologiques ont montré l'existence de phénomènes d'habituation, de curiosité, de recherche de sensations et de nouveauté, ainsi que l'existence de grandes capacités cognitives de réflexion, de mémoire et de compréhension. En fonction de tous ces facteurs, il est très probable qu'après plusieurs milliers d'explorations, d'expérimentations et de recherche de nouveaux plaisirs érotiques, les premiers être humains auraient progressivement découvert des activités érotiques de plus en plus élaborés (caresses érotiques, masturbation, masturbation réciproque, fellation, cunnilingus ...), jusqu'aux plus complexes (oro-génitale réciproque, coït vaginal, coït anal ...). Ensuite, la complémentarité morphologique du pénis et du vagin, la nature hautement érogène du vagin et sa lubrification, les sensations érotiques péniennes intenses provoqués par la pénétration, ainsi que la possibilité de maximaliser les stimulations somesthésiques et érotiques par d'autres activités simultanées (contact cutané d'une grande partie du corps, caresses, étreintes, baiser, ...), feraient que le coït vaginal deviendrait une des activités érotiques préférées.
Cette explication de la découverte et de l'apprentissage du coït vaginal est en grande partie spéculative. En raison de l'impossibilité de connaître le contexte des premiers Hommes, il est impossible de la vérifier. Néanmoins, au moins au niveau théorique, aucune donnée connue ne permet d'invalider cette explication. La découverte du coït vaginal semble possible, même en l'absence de toute information et connaissance, essentiellement grâce à la nature intensément érogène du pénis/clitoris et du vagin et grâce à leur complémentarité morphologique.
Remarques importantes & Généralisation
Ce modèle, chez l'Homme, de l'apprentissage du comportement permettant la reproduction, dont surtout de la séquence cruciale du coït vaginal, ne se situe pas dans le cadre de l' "instinct" et de la téléonomie, mais des paradigmes du « bricolage de l'évolution », de l'interactionnisme, de l'émergentisme (Figure 35) et du constructivisme. Ce serait à partir d'éléments innés, non optimisés par l'évolution, interagissant entre-eux et avec des caractéristiques de l'environnement, qu'émergerait un comportement de stimulation du corps. Ce qui serait inné ne seraient que des potentialités élémentaires et globales (détection de phéromones, réflexes, renforcement ...), qui néanmoins permettraient, mais uniquement de manière approximative, la réalisation des fonctions fondamentales, dont celle de la reproduction. Les schèmes moteurs du comportement érotique – dont ceux du coït vaginal –, les affects érotiques, les représentations et les valeurs sexuelles seraient acquis et se construiraient graduellement au cours du temps, principalement au cours de la période du développement.
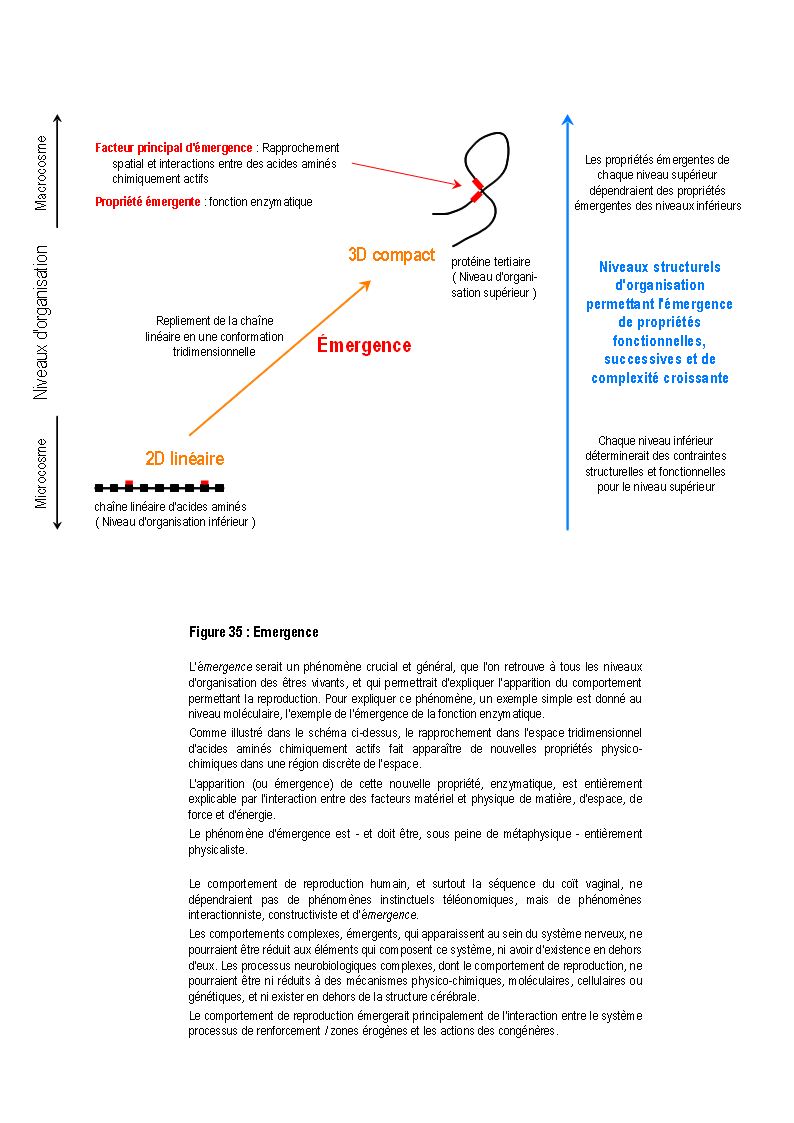
La généralisation des résultats de cette étude à l'ensemble des comportements humains suggère qu'il n'existerait pas d' "instincts" ou de "programmations" innés des comportements, mais plutôt tout un ensemble de processus neurobiologiques, innés mais élémentaires, à l'origine uniquement de tendances globales et approximatives, qui, au cours du développement et de l'interaction avec l'environnement, permettraient l'apprentissage par essais et erreurs de comportements relativement adaptés et appropriés à la survie de l'individu et de l'espèce.
Par exemple les deux tendances psychiques et innées de curiosité et de peur permettent une exploration approximative de l'environnement, avec parfois des comportements acquis inadaptés (peurs irrationnelles, curiosités inappropriés). Néanmoins ces deux tendances opposées sont suffisantes pour assurer l'exploration relativement adaptée de l'environnement et ainsi permettre la survie d'une majorité d'individus.
Si l'image – qui découle de ces cadres explicatifs – celle d'un être humain imparfait, principalement organisé pour des activités sexuelles, alimentaires, exploratoires, excrétoires et de sommeil, n'est pas aujourd'hui culturellement "noble" et "valorisante", elle semble néanmoins correspondre aux hasards de l'évolution et à une réalité biologique très pragmatique de nécessité impérieuse à réaliser, même approximativement, les fonctions fondamentales à la survie de l'individu et de l'espèce.
d – Discussion relative à la validité de l'hypothèse et du modèle
Après avoir présenté dans les chapitres précédants les analyses, les résultats, les problèmes et les limitations de cette recherche, quelle évaluation peut-on faire de la validité des conclusions finales ?
a – Validité de l'hypothèse principale
Par rapport à l'évaluation de la validité de l'hypothèse principale (l'acquisition chez l'Homme du comportement permettant la reproduction dépend – principalement mais indirectement – de l'activation des processus de renforcement, provoquée par la stimulation du corps et des zones érogènes des organes génitaux), on dispose des données suivantes :
– L'analyse préalable de la littérature a montré qu'il est probable qu'il n'existe pas d'instinct de la reproduction chez l'Homme (voir section "1.d – Le comportement de reproduction est-il inné ?", page 13). Ce résultat implique que ce comportement est très certainement appris.
– La recherche de la nature de cet apprentissage a montré qu'il est très probable que ce soient les processus de renforcement qui sont impliqués (voir section "1.e – Apprentissage du comportement permettant la reproduction", page 45).
– La vérification expérimentale, au moyen d'un questionnaire, a montré que la plupart des activités érotiques étaient centrées sur les organes génitaux, en raison de l'intensité du plaisir provoqué par la stimulation par un partenaire du pénis/clitoris et du vagin (voir chapitre "5.a – Discussion relative aux résultats expérimentaux", page 119). Ce résultat corrobore l'importance fonctionnelle du système constitué par les processus de renforcement et le pénis/clitoris/vagin.
– D'autres données, présentées dans la section suivante, qui corroborent l'ensemble du modèle, corroborent indirectement l'hypothèse principale.
– On ne connaît pas d'autres hypothèses ou modèles qui soient aussi satisfaisants (voir section "1.c – Théories actuelles et limites de ces théories", page 8).
– Enfin, aucune donnée actuellement connue ne réfute cette hypothèse.
Au terme de cette évaluation, et sous toutes réserves, tous ces éléments corroborent l'hypothèse.
b – Validité du modèle
La vérification systématique du modèle qui découle de l'hypothèse principale (voir section "2.b – Présentation détaillée du modèle comportemental", page 51), n'est pas l'objet de ce travail de recherche. Les éléments de corroboration présentés ci-dessous sont essentiellement destinés à conforter, bien qu'indirectement, l'hypothèse principale. Néanmoins, ces éléments, à savoir l'expérimentation ayant servie à la vérification de l'hypothèse principale, ainsi que des données recueillies au cours de cette étude, permettent dans le même temps de réaliser une évaluation globale du modèle théorique.
b.1 – Principales réfutations du modèle
Dans un premier temps, existe-t-il des faits objectifs qui réfuteraient le modèle ?
L'objection principale à ce modèle est le rôle majeur, expérimentalement vérifié, des hormones sexuelles et des phéromones dans la reproduction et le comportement sexuel. Bien qu'on observe que l'influence des hormones diminue avec le degré de développement du cortex, tant pour le comportement sexuel (BUVAT 1996) que d'ailleurs pour le comportement maternel (STERN 1997), il existe de nombreuses données qui montrent que la différenciation sexuelle de l'organisme et du système nerveux a encore une influence sur le comportement et l'orientation sexuelle des hommes et des femmes (MORRIS & al. 2004). Néanmoins, il convient de noter que le rôle primordial des renforcements érotiques n'est absolument pas contradictoire et incompatible avec un effet hormonal ou phéromonal. Ce sont des systèmes qui ont un fonctionnement complémentaire, et suivant la dynamique de mise en œuvre de ces systèmes, et suivant l'importance d'autres facteurs (émotionnels, cognitifs, environnementaux, culturels...), l'interactions de tous ces facteurs aboutira à un résultat comportemental singulier qui sera propre à chaque sujet. Le modèle de l'apprentissage du comportement permettant la reproduction n'exclut pas des effets hormonaux ou phéromonaux, il suppose uniquement que ces effets seraient, comparés aux effets majeurs des processus de renforcements érotiques, relativement plus faibles.
Par ailleurs, d'après le modèle théorique, la grande majorité des personnes devraient avoir des activités bisexuelles. Or, on observe dans les sociétés occidentales actuelles une sexualité quasi hétérosexuelle, ce qui, a priori, réfute le modèle. Néanmoins, on observe que quasiment tous les primates ont des activités bisexuelles (WALLEN & PARSONS 1997), en particulier les chimpanzés pan paniscus (Bonobo) (DE WALL 1992), que dans les sociétés sexuellement libérales les enfants et les adolescents ont des activités bisexuelles (FORD & BEACH 1965 ; MALINOWSKI 1970 ; DIAMOND 2004), et qu'apparemment il existait dans toutes les sociétés anciennes de guerriers, avant l'avènement des religions actuelles qui sont peu favorables à la sexualité, des pratiques bisexuelles généralisées (SERGENT 1986). Toutes ces données suggèrent qu'il existe une tendance significative à la bisexualité chez l'être humain. De plus, il faut prendre en compte en Occident la grande valorisation culturelle du couple hétérosexuel, une très forte homophobie (BAGLEY & TREMBLAY 2000), le fait que les bisexuels sont souvent rejetés par les hétérosexuels et également par les homosexuels, que la bisexualité n'existe pas au niveau des pratiques et des valeurs culturelles (RODRIGUEZ-RUST 2002), et qu'il est donc extrêmement difficile de vivre de manière bisexuelle (EVANS 2003). Afin de comprendre l'effet majeur de la pression de conformité et du contexte culturel, on peut donner comme premier exemple les normes sociales vestimentaires. Bien qu'il n'existe pas de lois ou d'interdits formels et que les personnes sont a priori "libres", on observe que la quasi totalité des hommes ne portent jamais d'habits de femmes. Tous les hommes se conforment aux codes implicites de la masculinité (BADINTER 1992). Cet exemple, relativement proche du domaine de la sexualité tout en ne dépendant d'aucun facteur biologique, permet de comprendre la puissance des normes sociales dominantes. Dans un autre registre, on peut donner l'exemple des conditionnements culturels alimentaires : dans les sociétés occidentales, on ne consomme pas d'annélides ou d'insectes, malgré leurs intérêts gustatifs et nutritionnels (DeFOLIART 1992), et l'idée d'en manger provoque en général du dégoût. Dans d'autres sociétés, les personnes sont par exemple prêtes à effectuer une journée de marche pour trouver des vers de palmier, et quand elles les mangent, on observe sur leur visage tous les signes du plaisir et du contentement. Enfin, encore dans un autre registre, on remarque que les personnes de certaines sociétés peuvent se suicider après être accidentellement entrées dans un cimetière, en raison de conditionnements émotionnels relatifs aux malédictions que les esprits des morts leurs auraient jeté. Ces trois exemples différents mettent en évidence l'importance majeure du facteur culturel – et surtout des conditionnements émotionnels concomitants – dans les comportements et les réactions affectives, et devraient permettre de comprendre l'effet vraisemblablement déterminant de l'homophobie et de l'hétérocentrisme sur le comportement et les affects sexuels. Malgré tout cela, on observe quand même qu'entre un tiers et la moitié des personnes occidentales ont eu au moins une expérience bisexuelle (KINSEY 1948), mais que vraisemblablement la plupart des personnes, en raisons de toutes les difficultés et pressions psychologiques exposées précédemment, se conforment aux pratiques et aux valeurs dominantes.
Une autre objection fréquente aux modèles différents du modèle dominant de l'hétérosexualité est la mise en évidence de caractéristiques particulières chez les homosexuels (par exemple LANDOLT & al. 2004), qui suggèrent que cet état (ainsi que les orientations autres qu'hétérosexuelles) ne serait pas "normal". Néanmoins, que ce soit pour la sexualité ou pour tout autre pratique, en raison de l'existence de très fortes pressions de conformité et de franche hostilité à la différence (cf. les paragraphes ci-dessus), il est très probable qu'une grande partie des caractéristiques particulières observées soient plus une conséquence qu'une cause du choix homosexuel (ou bisexuel).
Une autre réfutation possible serait apparemment l'existence d'une orientation sexuelle innée, mise en en évidence chez les homosexuels masculins, et dont l'origine serait due aux caractéristiques anatomiques et fonctionnelles de l'aire préoptique médiane (LEVAY 1991 ; SAVIC & al. 2005 ; BERGLUND & al. 2006). Néanmoins, même si ces résultats – bien qu'ils ne permettent pas actuellement de savoir si ces caractéristiques fonctionnelles sont innées ou acquises – correspondent effectivement à une orientation sexuelle inné, cela ne réfute pas le modèle. En effet, le modèle suppose uniquement la prépondérance des processus de renforcement érotique. Les effets des hormones et des phéromones peuvent exister, mais seraient plus faibles.
Enfin, certains résultats concernant les préférences pour les activités érotiques (cf. § 4.b.b, p. 91) semblent réfuter le modèle théorique : comment expliquer les préférences importantes pour des activités qui ne stimulent aucune zone érogène du sujet, telles par exemple pratiquer une fellation/cunnilingus ou masturber le sexe du partenaire ? En effet, lorsqu'une personne masturbe sont partenaire, il n'existe aucune relation anatomique directe entre les processus de renforcement du sujet et le pénis/clitoris du partenaire ; il n'existe donc aucun renforcement direct entre l'action de la main du sujet et les sensations érotiques provenant du sexe du partenaire. En fonction du modèle théorique, seul le partenaire devrait ressentir du plaisir érotique et le sujet ne devrait avoir aucune motivation à réaliser cet acte. Vraisemblablement, cet effet est secondaire et serait induit par certains processus émotionnels et cognitifs : conditionnement, association, empathie, mémorisation, anticipation ... Initialement, les personnes ne rechercheraient que les activités qui provoquent un renforcement érotique direct : masturbation par le sujet lui-même et stimulation de ses zones érogènes par un partenaire. Puis, graduellement, les activités érotiques effectuées par le sujet sur ses partenaires deviendraient également appétitives, vraisemblablement de la manière suivante. Dans un premier temps, on observe que déjà chez les rongeurs un stimuli même aversif peut devenir appétitif quand il est couplé à la copulation (PFAUS & al. 2001), ce qui montre la puissance des renforcements érotiques. Comme nous avons vu dans cette étude que le plaisir érotique procuré par un partenaire est le plaisir le plus intense que l'organisme humain peut ressentir, ce qui suggère l'importance fonctionnelle des renforcements érotiques chez l'Homme, il est donc vraisemblable que l'effet mis en évidence par Pfaus et ses collaborateurs chez les rongeurs existe également dans l'espèce humaine. Dans un second temps, la répétition des diverses activités érotiques entraîneraient également leur mémorisation et la mémorisation associée des états affectifs érotiques. De plus, des conditionnements de type pavlovien peuvent provoquer le rappel des états érotiques antérieurs juste par la perception d'un stimulus conditionné (vérifié expérimentalement chez le lapin, cité par FORD & BEACH 1965, p. 152). La capacité d'empathie (FRITH & FRITH 1999) peut induire chez le sujet un état mental similaire à celui qu'il observe ou déduit chez son partenaire. Les réactions positives du partenaire peuvent également être associées de manière gratifiante à l'activité érotique que réalise le sujet. Toutes ces caractéristiques émotionnelles et cognitives seraient à l'origine, progressivement, au cours du temps et de la répétition des activités érotiques, d'une association étroite entre, d'une part, les activités et les états érotiques du sujet, et, d'autre part, les activités et les états érotiques similaires de ses partenaires. C'est ce que déclare une des participantes à l'enquête, dans une remarque qui résume l'essentiel de cette interprétation : «Au début, je n'aimais pas faire les fellations demandées par mon compagnon, mais j'ai fini par aimer ça». Cette interprétation, partiellement étayée, nécessite néanmoins des vérifications complémentaires afin de préciser exactement les processus neurobiologiques à l'œuvre.
b.2 – Méthode systématique d'évaluation du modèle
Existe-t-il une méthode qui permettrait d'évaluer l'ensemble d'un modèle théorique d'une manière relativement fiable ?
À partir des travaux d'épistémologues et de philosophes des sciences (POPPER 1990, LADRIÈRE 1995, GIL 1995, LARGEAULT 1995 & 1999, CHALMERS 1982), il serait possible d'établir une méthode et une liste de critères qui permettraient d'évaluer globalement le « degré de crédibilité » (d'après Watanabe S. in LADRIÈRE 1995) d'une hypothèse ou d'une théorie.
Les critères d'évaluation de cette méthode seraient les suivants :
· Critères généraux :
– L'absence, après une recherche systématique qui tend à l'exhaustivité, de réfutation.
– La simplicité
– La minimalisation des spéculations.
– La cohérence interne, externe et factuelle.
– l'explicabilité et la prédictibilité
– La maximalisation des corroborations, par des facteurs et des données pluridisciplinaires.
· Critères biologiques :
– Phylogenèse : cohérence avec les connaissances phylogénétiques.
– Ontogenèse : cohérence avec les connaissances ontogénétiques.
· Conclusion :
– Tout ces critères devraient permettre d'évaluer le « degré de crédibilité » de la théorie.
La réfutation
Le critère de la réfutation (POPPER 1990) suppose qu'en l'absence de réfutation d'une hypothèse ou d'une théorie, plus cette réfutation a été recherché de manière systématique et exhaustive, plus le « degré de crédibilité » de cette hypothèse ou théorie est élevé.
Ce critère de validité est d'une importance majeure.
Or dans cette étude, malgré une expérimentation spécifique, l'analyse systématique de la littérature et la confrontation avec la diversité ethnologique des pratiques sexuelles, aucune donnée actuellement connue ne réfute ni l'hypothèse principale, ni apparemment le modèle théorique.
Simplicité
Le critère de simplicité suppose que plus une hypothèse ou une théorie est simple, plus son « degré de crédibilité » est élevé.
Néanmoins ce critère de validité n'est pas un critère important et déterminant. La simplicité est fréquente mais pas systématique dans les structures biologiques.
Par rapport à ce critère de simplicité, le modèle du comportement permettant la reproduction chez les primates hominoïdes est biologiquement simple. Il suffit de quelques éléments anatomiques et neurophysiologiques (processus de renforcement, pénis/clitoris ...) – qui existent déjà chez tous les mammifères, mais dont juste l'importance relative change chez les anthropoïdes – pour que l'acquisition du comportement de reproduction devienne hautement probable. Il n'est nul besoin de faire appel à des hypothèses sophistiquées de "programmation" spécifique, de "représentations préformées" ou de comportement de reproduction instinctuel précablé, qui nécessiteraient des organisations complexes au sein du système nerveux.
Minimalisation des spéculations
Ce critère suppose que moins il existe d'éléments spéculatifs dans une hypothèse ou une théorie, plus son « degré de crédibilité » est élevé.
Or aucun des principaux processus ou caractéristiques sur lesquels sont basés l'hypothèse et le modèle théorique de cette étude ne sont spéculatifs : les zones érogènes, les processus de renforcement, les organes génitaux intensément érogènes, les processus émotionnels et cognitifs, la conservation phylogénétique des principaux facteurs impliqués dans le comportement de reproduction, les modifications structurelles et fonctionnelles du système nerveux mammalien des rongeurs à l'Homme, l'importance de la stimulation corporelle dans la sexualité, la grande diversité des pratiques sexuelles, etc., sont connus ou démontrés expérimentalement, et sont acceptés par la communauté scientifique.
Cohérence interne
Ce critère suppose que plus une hypothèse ou une théorie possède une bonne cohérence interne, plus son « degré de crédibilité » est élevé.
Or l'hypothèse et le modèle proposés ne présentent pas d'incohérences, tant au niveau structurel que fonctionnel, et ne nécessitent aucune hypothèse supplémentaire ad hoc, nécessaire pour compléter le modèle afin de pouvoir expliquer tous les comportements sexuels (masturbation, activités homosexuelles, baiser, etc.).
Cohérence externe
Le critère de cohérence externe suppose que plus une hypothèse ou une théorie est en accord avec plusieurs autres hypothèses, théories ou connaissances déjà éprouvées, plus son « degré de crédibilité » est élevé.
Or l'hypothèse de cette étude est en accord avec les théories du renforcement en biologie et les théories de la récompense (Reward) en psychologie. De plus, ce nouveau paradigme de la sexualité humaine est en parfaite cohérence avec la théorie de l'évolution de François JACOB (1977, 1981).
Par ailleurs, l'hypothèse de l'apprentissage du comportement permettant la reproduction est conforme aux données connues relatives à l'importance majeure du système somatosensoriel dans le développement physiologique et psychique, à l'importance majeure du plaisir dans l'économie psychique (CABANAC 1971 ; 1992), à l'existence des réflexes moteurs sexuels (par exemple les poussées pelviennes, la lordose ...), à la diminution de l'influence des hormones sexuelles en fonction de la complexité cérébrale, à l'influence surtout physiologique des phéromones chez l'Homme, et enfin, au rôle des processus cognitifs, qui rendent possible l'élaboration du concept culturel de "sexualité" (constitué de représentations, de valeurs et de symbolisations complexes, qui influencent à la fois le développement du comportement "sexuel" et la perception que le sujet aura de tout ce qui est lié à la sphère définie comme "sexuelle").
Cohérence factuelle
Le critère de cohérence factuelle suppose que plus une hypothèse ou une théorie est en accord avec les faits, plus son « degré de crédibilité » est élevé.
Ce critère de validation est un critère majeur. Il est en effet crucial qu'une hypothèse soit conforme à la majorité, voire à la quasi totalité des faits.
Or le modèle présenté dans cette étude est conforme aux observations éthologiques des primates les plus proches de l'Homme et aux observations de la diversité des comportements sexuels humains, tant au niveau individuel qu'au niveau ethnologique dans les centaines de sociétés connues (FORD & BEACH 1965).
Enfin le modèle est cohérent avec les principales caractéristiques du comportement sexuel du chimpanzé pan paniscus (Bonobo). Ce fait est d'autant plus significatif que cette espèce serait la plus proche de l'Homme, tant au niveau génétique que cognitif et comportemental, et qu'il est difficile de supposer que son comportement sexuel puisse être "dénaturé" par l'influence culturelle. En particulier, la sexualité des chimpanzés pan paniscus démontre bien qu'un comportement dont le but est la stimulation du corps et non la procréation, permet néanmoins sans aucun problème particulier d'assurer la fonction fondamentale de la reproduction.
Phylogenèse
Le critère de la cohérence avec les connaissances phylogénétiques suppose que plus une hypothèse ou une théorie est en accord avec les données de la phylogenèse, plus son « degré de crédibilité » est élevé.
Or le modèle présenté est bien en accord avec la conservation au cours de l'évolution des principales molécules, structures et fonctions au sein du système nerveux mammalien. Il n'est nul besoin de faire disparaître, ou surtout faire apparaître, des systèmes fonctionnels. Les mêmes structures et fonctions, mais ayant évoluées, permettent d'expliquer les modifications de la dynamique comportementale.
Ontogenèse
Le critère de la cohérence avec les connaissances ontogénétiques suppose que plus une hypothèse ou une théorie est en accord avec les données de l'ontogenèse, plus son « degré de crédibilité » est élevé.
Or le modèle présenté est bien en accord avec le développement des structures et fonctions nécessaires à la dynamique fonctionnelle du modèle. Principalement, le système somatosensoriel, les réflexes sexuels et les processus de renforcement sont fonctionnels avant la naissance, tandis que la maturation de la coordination motrice intervient vers la troisième année, âge où peut apparaître, si le contexte est favorable, le comportement érotique.
L'explicabilité et la prédictibilité
Le critère d'explicabilité et de prédictibilité suppose que plus une hypothèse ou une théorie peut expliquer et prévoir les faits, plus son « degré de crédibilité » est élevé.
Ce critère de validité est d'une importance majeure.
Or le modèle présenté dans cette étude permet de comprendre et d'expliquer toute la diversité des activités érotiques humaines, et de plus à partir d'un seul et unique modèle. Les grands types d'activités érotiques connus (masturbation, activités homoérotiques ou hétéroérotiques, en couple ou en groupe, activités érotiques avec des animaux ou avec des objets), ayant actuellement chacun une explication causale différente (reproduction, instinct, plaisir, pathologie, perversions...), peuvent être regroupés dans un seul modèle neurobiologique explicatif, global et cohérent. Ces activités deviennent alors "logiques", "cohérentes" et "biologiquement normales" (même si dans certaines sociétés, certaines de ces pratiques peuvent être "culturellement anormales"). En effet, si le plaisir physique intense, principalement provoqué par des stimulations mécaniques, est bien le facteur neurobiologique majeur à l'origine des activités érotiques, il est biologiquement "logique" et "normal" que l'être humain cherche à reproduire toutes les situations de jouissance érotique et orgastique qu'il découvre, quel qu’en soit le moyen : autostimulation, un homme ou une femme, un ou plusieurs partenaires, un animal, un objet, ou bien – moyen artificiel mais le plus efficace et direct pour stimuler les régions cérébrales du plaisir – l'injection d'une molécule psycho-active érogène.
De plus, ce modèle permet d'expliquer et de prévoir les principales modifications du comportement sexuel dans chaque espèce (ou plutôt pour chaque ordre) de mammifère, en fonction de l'importance relative de chaque facteur de la reproduction (réflexes sexuels, hormones, phéromones, processus de renforcement).
Maximalisation des corroborations
Le critère de maximalisation des corroborations suppose, qu'en l'absence de réfutation, plus il existe d'éléments et de données qui confortent une hypothèse ou une théorie, plus son « degré de crédibilité » est élevé.
Ce critère de validation est un critère important.
En rapport à ce critère, la majorité des données pluridisciplinaires (génétique, biologie moléculaire, neuroanatomie-fonctionnelle, imagerie cérébrale, éthologie, ethnologie, psychologie…), recueillis par une analyse de la littérature, des observations, des interviews et des enquêtes, et présentées tout au long de cette étude, confortent l'hypothèse.
De plus, la conformité de l'hypothèse et du modèle aux différents critères présentés dans les paragraphes ci-dessus est un élément supplémentaire de corroboration.
e – Conclusion relative à la validité de l'hypothèse et du modèle
En fonction de toutes les analyses réalisées dans ce chapitre "Discussion", quelle est alors la validité de l'hypothèse et du modèle théorique ?
Aucune donné actuellement disponible ne permet d'affirmer avec une absolue certitude que l'hypothèse et le modèle proposés rendent compte de la réalité des processus biologiques à l'œuvre chez l'être humain.
Néanmoins, comme il n'existe pas de données connues qui réfutent cette hypothèse, et qu'il existe beaucoup de données et d'éléments qui la corrobore, on peut donc formuler la conclusion suivante :
L'hypothèse de l'importance primordiale des processus de renforcement, ainsi que le modèle global de l'apprentissage du comportement érotique qui en découle, seraient, à partir des différentes données scientifiques actuellement connues et disponibles, le moins mauvais modèle qu'il est aujourd'hui possible de proposer et qui permette d'expliquer l'origine et la dynamique du comportement permettant la reproduction, ainsi que de préciser les fondements primordiaux de la sexualité humaine.
|
Conclusion générale
L'objectif principal de ce travail de recherche était d'identifier et de comprendre la dynamique des différents facteurs biologiques fondamentaux et primordiaux qui sont à l'origine du comportement permettant la reproduction, essentiel à la survie de l'espèce humaine.
Principal résultat
Dans la première partie de cette recherche, nous avons vérifié à partir de l'analyse des données éthologiques, ethnologiques et neurobiologiques, que, chez l'Homme, le comportement de reproduction (dont tout particulièrement la séquence cruciale du coït vaginal) ne serait pas inné.
Puis nous avons vérifié, en partie au moyen d'une expérimentation, que, chez l'Homme, l'acquisition du comportement permettant la reproduction dépendrait – principalement mais indirectement – de l'activation des processus de renforcement, provoquée par la stimulation du corps et des zones érogènes des organes génitaux.
Cette double vérification, montrer que le comportement permettant la reproduction n'est pas inné et qu'il est acquis, est nécessaire, car le fait de démontrer l'existence d'apprentissages ne permet pas d'exclure l'hypothèse de l'innéité de ce comportement. En effet, il est toujours possible d'objecter que les apprentissages ont masqué les processus innés, et que, s'il n'y avait pas eu d'apprentissages, ces processus innés auraient été révélés et auraient permis la réalisation du comportement de reproduction.
Cette double vérification nous permet de conclure, en résumant l'essentiel, que, chez l'Homme, le comportement permettant la reproduction ne serait pas inné, mais acquis.
Le « bricolage de l'évolution »
Pour bien comprendre les résultats de cette recherche, il est nécessaire de les interpréter dans le cadre du paradigme du « bricolage de l'évolution » de François Jacob (1977 ; 1981). Contrairement au modèle dominant de la théorie néodarwinienne qui suppose une optimisation des organismes qui tend presque à la perfection, il semblerait que l'effet majeur de la sélection naturelle n'est pas tant cette optimisation que l'élimination des organismes non viables. Ce qui signifie que les organismes actuels ne sont pas forcément parfaits ni même optimisés, mais que leurs structures anatomiques et physiologiques leur permet – peut importe la manière – de survivre et de se reproduire.
Facteurs primordiaux à l'origine du comportement permettant la reproduction
A l'issue de toutes les analyses présentées dans cette étude, quels sont les facteurs à l'origine de l'apprentissage du coït vaginal ?
Le premier facteur inné, primordial et principal, serait un système fonctionnel constitué par les processus de renforcement (localisés dans la partie basse du faisceau médian du télencéphale – noyau accumbens, aire tegmentale ventrale ...), associés au système somatosensoriel (et en particulier aux zones érogènes mucocutanées orales et pelviennes, dont principalement le pénis /clitoris ). Des processus hédoniques (localisés dans la partie haute du faisceau médian du télencéphale – septum latéral, lobe préfrontal ventromédian ...) seraient également étroitement associés à ce système fonctionnel. Cette organisation structurelle provoquerait la répétition de la stimulation du corps et des organes génitaux, ainsi que des sensations conscientes de plaisirs intenses associées à ces stimulations (plaisir sensuels et érotiques, orgasme).
Le deuxième facteur inné, primordial mais plus secondaire, serait un système hormonal et phéromonal (principalement localisé au niveau hypothalamique – aire préoptique médiale, hypothalamus antérieur ...). Cette organisation structurelle et moléculaire provoquerait à la puberté une augmentation significative du désir et des activités sexuelles, ainsi qu'éventuellement une tendance – mais faible – au rapprochement hétérosexuel, sous l'effet des phéromones.
La structure (neuro)biologique principale – processus de renforcement associés aux zones érogènes (pénis/clitoris principalement) – similaire aux deux sexes, crée les conditions d'un apprentissage hautement probable d'une grande variété de séquences motrices érotiques, dont certaines, telles le coït vaginal fécondant, deviennent préférentielles. La grande intensité du renforcement érotique serait à l'origine d'une importante fréquence des activités érotiques, qui est constatée par exemple dans les sociétés où n'existent pas ou peu de restrictions culturelles à la sexualité. Cette fréquence élevée permet ainsi la réalisation d'un nombre plus que suffisant de coïts fécondants, et, par voie de conséquence, la reproduction et la survie de l'espèce.
Par rapport à la phase motivationnelle du comportement sexuel, c'est-à-dire au rapprochement des partenaires, les renforcements liés à la stimulation hédonique du corps (plaisirs somatosensoriels) représenteraient environ les deux tiers des facteurs à l'origine du désir d'avoir des activités érotiques avec un partenaire. Le tiers restant serait essentiellement constitué par des facteurs émotionnels et cognitifs (amour, tendresse, complicité, curiosité ...).
Par rapport à la phase consommatoire du comportement sexuel, c'est-à-dire à la réalisation effective du comportement érotique, les renforcements érotiques seraient le facteur majeur, et peut être unique, à l'origine de l'organisation des activités érotiques autour de 3 zones érogènes majeures : la zone génitale (principalement), et les zones buccale et anale (secondairement).
Comportement de reproduction, comportement érotique & Pan-sexualité potentielle
Toutes ces données suggèrent qu'il n'existerait pas chez l'Homme de comportement de reproduction inné, c'est-à-dire une organisation neurale hautement spécifique qui contrôle sans apprentissage préalable le rapprochement hétérosexuel des partenaires, ainsi que les séquences motrices du coït vaginal permettant la fécondation.
La prépondérance fonctionnelle d'un système constitué par les processus de renforcement et le système somatosensoriel serait à l'origine de l'apprentissage d'un comportement dont le but est la stimulation du corps, et non la reproduction. Le coït vaginal reproducteur serait ainsi une conséquence indirecte et presque fortuite de la recherche des plaisirs somatosensoriels. Ce comportement, dont le but perçu au niveau conscient est le plaisir intense, pourrait être appelé "comportement érotique ".
Le fait que toute stimulation corporelle qui est érotiquement renforcé sera répétée – si le contexte le permet – fait qu'il existe une grande potentialité d'activités qui peuvent devenir érotiques. L'existence de capacités cognitives développées rend possible la création potentielle du concept de "sexualité" et son attribution, également potentielle, à une grande variété d'activités, d'éléments psychiques et de situations. Ces caractéristiques font que la sexualité humaine serait acquise et aurait une potentialité pan-sexuelle.
Analyse fonctionnelle et phylogénétique
En synthèse, on observe que l' anatomie et la physiologie de la reproduction sont innées, tandis que le comportement permettant la reproduction est acquis. Cela signifie que ce qui est inné sont les conditions qui rendent hautement probable l'acquisition du comportement permettant la reproduction.
Chez l'Homme, ce qui est biologiquement organisé de manière innée, c'est essentiellement la stimulation potentielle du corps.
On observe qu'il s'agit d'une organisation minimale. Ce qui est inné est juste l'essentiel : la tendance à la stimulation préférentielle du pénis/clitoris (et éventuellement du vagin). Le reste, c'est-à-dire le détail des séquences motrices permettant les stimulations érotiques ainsi que le coït vaginal fécondant, est appris grâce aux processus émotionnels et cognitifs. Il est important de remarquer que ces processus émotionnels et cognitifs ne sont pas spécifiques à la sexualité. Ces processus participent également à l'apprentissage des autres comportements.
Au niveau phylogénétique, chez les mammifères, on observe que le contrôle du comportement sexuel devient moins moléculaire et se déplace vers le télencéphale : principalement localisé dans le mésencéphale et le diencéphale avec surtout des hormones et des phéromones chez les rongeurs, et principalement localisé dans le diencéphale et le télencéphale avec plutôt des processus émotionnels et cognitifs chez l'Homme.
Par ailleurs, l'analyse phylogénétique suggère l'existence initiale chez les proto-mammifères d'un comportement spécifiquement organisé pour la reproduction (inhibition saisonnière par la mélatonine, rut, hormones sexuelles, phéromones sexuelles, lordose, érection, poussées pelviennes, réflexe éjaculatoire, libération de l'ovule lors du coït ...) dont une partie des différents éléments constitutifs auraient été perdus ou modifiés lors des transformations du système nerveux au cours de l'évolution. Les éléments restant chez l'Homme, bien que profondément modifiés, permettent toujours la reproduction, mais d'une manière indirecte, particulière et contre-intuitive. C'est là un parfait exemple du « bricolage de l'évolution ».
Toutes ces caractéristiques phylogénétiques, ontogénétiques et psychobiologiques, qui ont été mise en évidence dans cette étude pour le comportement de reproduction (cf. Figure 23, page 55 ), existeraient également dans la dynamique fonctionnelle des autres comportements humains.
Affectivité & Sexualité
De manière plus générale, il semble exister un phénomène psychobiologique d'ordre "affectif", plus large que ce qui est aujourd'hui culturellement défini comme "sexualité". La stimulation du corps (chatouilles, contacts corporels, caresses sensuelles, stimulations érotiques) correspondrait au niveau le plus basique, le cœur primordial de ce phénomène. Puis s'ajouterait successivement les différents effets, immédiats puis ultérieurs, de cette stimulation : effets sensoriels, effets renforçants, effets d'attachement et parfois de dépendance, effets émotionnels et effets cognitifs.
Ce qui est culturellement défini comme étant la "sexualité" correspondrait en fait à une partie de ce phénomène biologique affectif, associé avec des représentations cognitives élaborées. L'ensemble des données disponibles suggère que la "sexualité" serait le résultat – à partir des stimulations répétées du corps et des effets émotionnels et passionnels que ces stimulations entraînent – d'une construction sensorielle, émotionnelle, cognitive et culturelle, tout au long du développement et de l'existence du sujet.
Bibliographie
ADAM Barry D. : Age, structure, and sexuality : reflections on the anthropological evidence on homosexual relations · in BLACKWOOD Evelyne (Ed) : The many faces of homosexuality : anthropological approaches to homosexual behavior · Harrington Park Press, New York, 1986
ADKINS-REGAN Elisabeth : Is the snark still a boojum ? The comparative approach to reproductive behavior · Neuroscience and biobehavioral reviews, 14:243-252, 1990
AGMO Anders , BERENFELD R. Reinforcing properties of ejaculation in the male rat : role of opioids and dopamine -- Behavioral Neuroscience, 104(1):177-182, 1990
AGMO Anders , ELLINGSEN Ellinor. Relevance of non-human animal studies to the understanding of human sexuality -- Scandinavian Journal of Psychology, 44(3):293-301, 2003
AGMO Anders , GOMEZ M. Sexual reinforcement is blocked by infusion of naloxone into the medial preoptic area -- Behavioral Neuroscience, 107(5):812-818, 1993
AGMO Anders , VILLALPANDO Adriana , PICKER Ziva , FERNANDEZ Hilda. Lesions of the medial prefrontal cortex and sexual behavior in the male rat -- Brain Research, 696(1-2):177-186, 1995
AGMO Anders. Copulation-contingent aversive conditioning and sexual incentive motivation in male rats : evidence for a two-stage process of sexual behavior -- Physiology & Behavior, 77(2-3):425-435, 2002
AGMO Anders. Lack of opioid or dopaminergic effects on unconditioned sexual incentive motivation in male rats -- Behavioral Neuroscience, 117(1):55-68, 2003
AGMO Anders. Male rat sexual behavior -- Brain Research : Brain Research Protocols, 1(2):203-209, 1997
AGMO Anders. Sexual motivation - an inquiry into events determining the occurrence of sexual behavior -- Behavioural Brain Research, 105(1):129-150, 1999
AGMO Anders. Unconditioned sexual incentive motivation in the male Norway rat (Rattus norvegicus) -- J. Comp Psychol., 117(1):3-14, 2003
ALEANDRI V. , SPINA V. , MORINI A. The pineal gland and reproduction -- Hum. Reprod. Update., 2(3):225-235, 1996
ALLARD J. , TRUITT W. A. , MCKENNA K. E. , COOLEN L. M. Spinal cord control of ejaculation -- World J. Urol., 23(2):119-126, 2005
ALLGEIER Albert Richard , ALLGEIER Elisabeth Rice : Sexualité humaine ? De Boeck Université 1992
ALONI Ronit , KATZ Shlomo. A review of the effect of traumatic brain injury on the human sexual response -- Brain Injury, 13(4):269-280, 1999
ALZATE H. Vaginal erogeneity, "female ejaculation," and the "Grafenberg spot" -- Archives of Sexual Behavior, 19(6):607-611, 1990
ALZATE H. Vaginal eroticism and female orgasm: a current appraisal -- Journal of Sex and Marital Therapy, 11(4):271-284, 1985
ALZATE H. Vaginal eroticism: a replication study -- Archives of Sexual Behavior, 14(6):529-537, 1985
AMINI Fariborz , LEWIS Thomas , LANNON Richard , LOUIE A. , BAUMBACHER G. , MCGUINNESS T. , SCHIFF E. Z. Affect, attachment, memory: contributions toward psychobiologic integration -- Psychiatry, 59(3):213-239, 1996
ANAPOL Deborah M. : Polyamory : the new love without limits ? Intinet ressource center 1997
ANDERSON K.A. , WAGNER G. : Physiology of penile erection ? Physiological reviews, 75:191-235, 1995
ARIES Philippe : Le mariage indissoluble ? Communications, 35:123-137, 1982
ARIES Philippe : Saint Paul et la chair ? Communications, 35:34-36, 1982
ARNOW B. A. , DESMOND J. E. , BANNER L. L. , GLOVER G. H. , SOLOMON A. , POLAN M. L. , LUE T. F. , ATLAS S. W. Brain activation and sexual arousal in healthy, heterosexual males -- Brain, 125(Pt 5):1014-1023, 2002
ARON Claude : La bisexualité et l'ordre de la nature ? Odile Jacob 1996
ARRINGTON R. , COFRANCESCO J. , WU A. W. Questionnaires to measure sexual quality of life -- Quality of Life Research, 13(10):1643-1658, 2004
BADINTER Elisabeth : XY, de l'identié masculine · Odile Jacob, Poche, 1992
BADINTER Elisabeth : L'amour en plus · Flammarion Poche 1980
BAGATELL CJ , HEINMAN JR , RIVIER JE , BREMNER WJ : Effects of testosterone and estradiol on sexual behavior in normal young men · The journal of clinical endocrinology and metabolism, 78:711-716, 1994
BAGEMIHL Bruce : Biological exuberance ? St Martin Press, New York, 2000
BAGLEY C. , TREMBLAY P. Elevated rates of suicidal behavior in gay, lesbian, and bisexual youth -- Crisis, 21(3):111-117, 2000
BAGLEY C. , TREMBLAY P. Suicidal behaviors in homosexual and bisexual males -- Crisis, 18(1):24-34, 1997
BALTHAZART J. , FABRE-NYS C. Le comportement sexuel -- in THIBAULT C. , LEVASSEUR M.-C. (Eds). La reproduction chez les mammifères et l'Homme -- INRA Ellipse, 2(27):611-637, 2001
BANCROFT J. The endocrinology of sexual arousal -- J. Endocrinol., 186(3):411-427, 2005
BANCROFT John , EVERITT Barry J. Of rats and men : the comparative approach to male sexuality -- Annual Review of Sex Research, 2:77-117, 1991
BANCROFT John , JANSSEN Erik. The dual control model of male sexual response: a theoretical approach to centrally mediated erectile dysfunction -- Neuroscience and Biobehavioral Reviews, 24(5):571-579, 2000
BANCROFT John. Biological factors in human sexuality -- The Journal of Sex Research, 39(1):15-21, 2002
BANCROFT John. Central inhibition of sexual response in the male : a theoretical perspective -- Neuroscience and Biobehavioral Reviews, 23(6):763-784, 1999
BARFIELD RJ, AUERBACH P, GEYER LA, McINTOSH TK. Ultrasonic vocalizations in rat sexual behavior. · Am Zool 1979;19:469±80
BARTELS Andreas , ZEKI Semir. The neural basis of romantic love -- Neuroreport, 11(17):3829-3834, 2000
BARTELS Andreas , ZEKI Semir. The neural correlates of maternal and romantic love -- Neuroimage, 21(3):1155-1166, 2004
BARTOSHUK Linda M. ; BEAUCHAMP Gary K. : Chemical sense ? Annual review of psychology, 45:419-449, 1994
BATESON Gregory P. P. Empreinte -- in GREGORY Richard L. (Ed). Le cerveau. Dictionnaire encyclopédique -- Robert Laffont, 373-376, 1993
BAUM M. J. , BRESSLER S. C. , DAUM M. C. , VEIGA C. A. , MCNAMEE C. S. Ferret mothers provide more anogenital licking to male offspring: possible contribution to psychosexual differentiation -- Physiology & Behavior, 60(2):353-359, 1996
BAUM Michael J. : Frank Beach's research on the sexual differentiation of behavior and his struggle with the "organizational" hypothesis ? Neuroscience and biobehavioral reviews, 14:201-206, 1990
BEATTY William W. : Gonadal hormones and sex differences in nonreproductive behaviors · in GERAL Arnold A. , MOLTZ Howard , WARD Ingeborg L. (Ed) : Sexual differentiation, vol 11, Handbook of behavioral neurobiology · Plenum Press, NY, 1992
BECKER Jill B., RUDICK Charles N., and JENKINS William J. The Role of Dopamine in the Nucleus Accumbens and Striatum during Sexual Behavior in the Female Rat. The Journal of Neuroscience, May 1, 21(9):3236-3241, 2001
BEJIN André : Crépuscule des psychanalyste, matin des sexologues · Communications, 35:159-177, 1982
BEJIN André : Le mariage extra conjugal · Communications, 35:138-146, 1982
BEJIN André : Le pouvoir des sexologues et la démocratie sexuelle · Communications, 35:178-192, 1982
BENATTAR Marie-Claude. L'amplification du point G, une nouvelle approche thérapeutique des dysfonctions sexuelles féminines ? -- Sexologies, XIV(51):5-10, 2005
BERGLUND H. , LINDSTROM P. , SAVIC I. Brain response to putative pheromones in lesbian women -- Proceedings of the National Academy of Sciences of the United States of America, 103(21):8269-8274, 2006
BERLINER D. L. , JENNINGS-WHITE C. , LAVKER R. M. The human skin: fragrances and pheromones -- J. Steroid Biochem. Mol. Biol., 39(4B):671-679, 1991
BERLINER David L. , MONTI-BLOCH Luis , JENNINGS-WHITE Clive , DIAZ-SANCHEZ Vincente : The functionality of the human vomeronasal organ (VNO) : evidence for steroid receptors ? Journal of steroid biochemistry and molecular biology, 58/3:259-265, 1996
BERRIDGE Kent C. Measuring hedonic impact in animals and infants: microstructure of affective taste reactivity patterns -- Neuroscience and Biobehavioral Reviews, 24(2):173-198, 2000
BESSON J.M. , GUILBAUD G. , BESSON J.M. : Peripheral neurons in nociception : physio-pharmacological aspects · John Libbey Eurotext, Paris, 1994
BIALY Michal , KACZMAREK Leszek : c-fos expression as a tool to search for the neurobiological base of the sexual behaviour of males ? Acta neurobiologiae experimentalis, 56:567-577, 1996
BINET André : L'amour et l'émotion chez la femme ? Vigot frères 1948
BLACKWOOD Evelyn : Breaking the mirror : the construction of lesbianism and the anthropological discourse on homosexuality ? in BLACKWOOD Evelyne (Ed) : The many faces of homosexuality : anthropological approaches to homosexual behavior ? Harrington Park Press, New York, 1986
BLOOD Anne J. , ZATORRE Robert J. Intensely pleasurable responses to music correlate with activity in brain regions implicated in reward and emotion -- Proceedings of the National Academy of Sciences of the United States of America, 98(20):11818-11823, 2001
BOCQUET Charles , MANTRAN Robert. Sélection naturelle -- Encyclopedia Universalis v. 11. 0, 2005
BOEHM N. : Environnement olfactif et reproduction : rôle de la voie olfactive accessoire ? Journal de médecine de Strasbourg, 23/3:138-143, 1992
BOEHM U. , ZOU Z. , BUCK L. B. Feedback loops link odor and pheromone signaling with reproduction -- Cell, 123(4):683-695, 2005
BOGAERT A. F. Asexuality: prevalence and associated factors in a national probability sample -- The Journal of Sex Research, 41(3):279-287, 2004
BONDIL Pierre , WESPES Eric : Anatomie et Physiologie de l'érection ? Progrès en urologie, 2/5:719-857, 1992
BOSSY Jean (Ed) : Anatomie clinique : neuroanatomie · Springer-Verlag 1990
BOURGEOIS Marc-Louis. L'anhédonie : le non-plaisir et la psychopathologie -- Masson, 1999
BRACKETT Nancy L. , FERRELL Sean M. , ABALLA Teodoro C. , AMADOR M. J. , PADRON O. F. , SONKSEN J. , LYNNE C. M. An analysis of 653 trials of penile vibratory stimulation in men with spinal cord injury -- The Journal of Urology, 159(6):1931-1934, 1998
BREGLIANO Jean-Claude. Variation génétique adaptative -- Encyclopedia Universalis v. 11. 0, 2005
BRENOT Philippe (Ed) – Dictionnaire de la sexualité humaine. L’Esprit du Temps, Bordeaux, 2004.
BRENOT Philippe : Éloge de la masturbation · 1997
BRENOT Philippe : L'éducation sexuelle · PUF QSJ 1996
BRODY S. , KRUGER T. H. The post-orgasmic prolactin increase following intercourse is greater than following masturbation and suggests greater satiety -- Biol. Psychol., 71(3):312-315, 2006
BROUSSIN Bernard , BRENOT Philippe. Orgasme in utero ? -- Sexologies, 21(5):15-16, 1996
BROUSSIN Bernard , BRENOT Philippe. Existe-t-il une sexualité du fœtus ? Fertilité, contraception, sexualité, Nov, 23/11:696-698, 1995
BRUCE A. , COHN M.D. : In search of human skin pheromones ? Arch. dermatol., 130:1048-1051, 1994
BUCK Linda B. Information coding in the vertebrate olfactory system -- Annu. Rev. Neurosci., 19:517-544, 1996
BULLOUGH Vern L. Alfred Kinsey and the Kinsey report : historical overview and lasting contribution -- The Journal of Sex Research, 35(2):127-131, 1998
BULLOUGH Vern L. Children and adolescents as sexual beings : a historical overview -- Child and Adolescent Psychiatric Clinics of North America, 13(3):447-59, 2004
BURGDORF Jeff , PANKSEPP Jaak. Tickling induces reward in adolescent rats -- Physiology & Behavior, 72(1-2):167-173, 2001
BURLESON M. H. , TREVATHAN W. R. , GREGORY W. L. Sexual behavior in lesbian and heterosexual women: relations with menstrual cycle phase and partner availability -- Psychoneuroendocrinology, 27(4):489-503, 2002
BURLESON Mary H. ; GREGORY Larry W. ; TREVATHAN Wenda R. : Heterosexual activity : relationship with ovarian function ? Psychoneuroendocrinology, 20/4:405-421, 1995
BURRIS A. S. , BANKS S. M. , CARTER C. S. , DAVIDSON J. M. , SHERINS R. J. A long-term, prospective study of the physiologic and behavioral effects of hormone replacement in untreated hypogonadal men -- Journal of Andrology, 13(4):297-304, 1992
BUSER Pierre. Motricité · Encyclopaedia Universalis, version CD-ROM 11.0, 2005
BUSTON Peter M. , EMLEN Stephen T. Cognitive processes underlying human mate choice : The relationship between self-perception and mate preference in Western society -- Proceedings of the National Academy of Sciences of the United States of America, 100(15):8805-8810, 2003
BUVAT J. , LEMAIRE A. , RATAJCZYK J. : Rôle des hormones dans les dysfonctions sexuelles, l'homosexualité, le transsexualisme et les comportements sexuels deviants : conséquence diagnostiques et thérapeutiques ? Contraception, fertilité, sexualité, 24/11:834-846, 1996
BUVAT J. : Hormones et comportement sexuel de l'Homme : données physiologiques et physiopathologiques ? Contracept. Fertil. Sex., 24/10:767-778, 1996
CABANAC M. Physiological role of pleasure -- Science, 173(2):1103-1107, 1971
CABANAC M. Pleasure: the common currency -- Journal of Theoretical Biology, 155(2):173-200, 1992
CABANAC Michel , GUILLAUME Jacqueline , BALASKO Marta , FLEURY Adriana. Pleasure in decision-making situations -- BMC. Psychiatry, 2(1):7, 2002
CAGGIULA A. R. Analysis of the copulation-reward properties of posterior hypothalamic stimulation in male rats -- J. Comp Physiol Psychol., 70(3):399-412, 1970
CAGGIULA A. R. , HOEBEL B. G. "Copulation-reward site" in the posterior hypothalamus -- Science, 153(741):1284-1285, 1966
CALLENDER Charles , Kochems Lee M. : Men and not-men : male gender-mixing statuses and homosexuality ? in BLACKWOOD Evelyne (Ed) : The many faces of homosexuality : anthropological approaches to homosexual behavior ? Harrington Park Press, New York, 1986
CAMI J. , FARRE M. Drug addiction -- N. Engl. J. Med., 349(10):975-986, 2003
CARDINAL Rudolf N. , EVERITT Barry J. Neural and psychological mechanisms underlying appetitive learning : links to drug addiction -- Current Opinion in Neurobiology, 14(2):156-162, 2004
CARDINAL Rudolf N. , PARKINSON John A. , HALL Jeremy , EVERITT Barry J. Emotion and motivation : the role of the amygdala, ventral striatum, and prefrontal cortex -- Neuroscience and Biobehavioral Reviews, 26(3):321-352, 2002
CARMICHAEL M. S. , WARBURTON V. L. , DIXEN J. , DAVIDSON J. M. Relationships among cardiovascular, muscular, and oxytocin responses during human sexual activity -- Archives of Sexual Behavior, 23(1):59-79, 1994
CARTER C. S. , DEVRIES A. C. , TAYMANS S. E. , ROBERTS R. L. , WILLIAMS J. R. , GETZ L. L. Peptides, steroids, and pair bonding -- Annals of the New York Academy of Sciences, 807:260-272, 1997
CARTER C. Sue , DEVRIES A. Courtney , GETZ Lowell L. Physiological substrates of mammalian monogamy: the prairie vole model -- Neuroscience and Biobehavioral Reviews, 19(2):303-314, 1995
CARTER C. Sue. Neuroendocrine perspectives on social attachment and love -- Psychoneuroendocrinology, 23(8):779-818, 1998
CASS A. S. , BLOOM Beth Ann , LUXENBERG M. Sexual fonction in adults with myelomeningocele -- The Journal of Urology, 136:425-426, 1986
CELA-CONDE C. J. , MARTY G. , MAESTU F. , ORTIZ T. , MUNAR E. , FERNANDEZ A. , ROCA M. , ROSSELLO J. , QUESNEY F. Activation of the prefrontal cortex in the human visual aesthetic perception -- Proceedings of the National Academy of Sciences of the United States of America, 101(16):6321-6325, 2004
CENTENO M. L. , LUO J. , LINDSTROM J. M. , CABA M. , PAU K. Y. Expression of alpha 4 and alpha 7 nicotinic receptors in the brainstem of female rabbits after coitus -- Brain Research, 1012(1-2):1-12, 2004
CHALMERS Alan F. : Qu'est-ce que la science ? · La Découverte (poche) 1982
CHANGEUX J. P. , DANCHIN A. Selective stabilisation of developing synapses as a mechanism for the specification of neuronal networks -- Nature, 264(5588):705-712, 1976
CHILTON J. K. Molecular mechanisms of axon guidance -- Dev. Biol., 292(1):13-24, 2006
CHIVERS M. L. , RIEGER G. , LATTY E. , BAILEY J. M. A sex difference in the specificity of sexual arousal -- Psychol. Sci., 15(11):736-744, 2004
COGLITORE Maria-Teresa. Bien-être et santé sexuelle -- Sexologies, XIII(49):45-48, 2004
COHEN Jonathan D. , BLUM Kenneth I. Reward and decision -- Neuron, 36(2):193-198, 2002
COHEN Jonathan D. , NOLL Douglas C. , SCHNEIDER Walter. Functional magnetic resonance imaging : overview and methods for psychological research -- Behavior Research Methods, Instruments & Computers, 25(2):101-113, 1993
COLAPINTO John. As nature made him. The boy who was raised as a girl. HarperCollins, 2000
COLD C. J. , MCGRATH K. A. Anatomy and histology of the penile and clitoral prepuce in primates -- in DENNISTON G. C. , HODGES F. M. , MILOS M. F. (Eds). Male and female circumcision -- Kluwer Academic / Plenum Publisher, 1999
COLD C. J. , TAYLOR J. R. The prepuce -- BJU. Int., 83 Suppl 1:34-44, 1999
COMFORT Alex : Les joies du sexe ? Arthaud 1992
CONSTANTINE (a) Larry L. : The effects of early sexual experiences : a review and synthesis of research ? in CONSTANTINE Larry L. , MARTINSON Floyd M. , (Ed) : Children and sex : new findings, new perspectives ? Little, Brown & Co., Boston, 1981
CONSTANTINE Larry L. , MARTINSON Floyd M. , (Ed) : Children and sex : new findings, new perspectives · Little, Brown and Co, Boston, 1981
COOKE B. M. , CHOWANADISAI W. , BREEDLOVE S. M. Post-weaning social isolation of male rats reduces the volume of the medial amygdala and leads to deficits in adult sexual behavior -- Behavioural Brain Research, 117(1-2):107-113, 2000
COOLEN L. M. Neural control of ejaculation -- J. Comp Neurol., 493(1):39-45, 2005
COSNIER Jacques : Psychologie des émotions et des sentiments · Retz 1994
COUREAUD G. , MONCOMBLE A. S. , MONTIGNY D. , DEWAS M. , PERRIER G. , SCHAAL B. A pheromone that rapidly promotes learning in the newborn -- Curr. Biol., 16(19):1956-1961, 2006
COWLEY J.J. , BROOKSBANK B.W. : Human exposure to putative pheromones and changes in aspects of social behavior ? Journal of steroid biochemistry and molecular biology, 39/4B:647-659, 1991
CRAWFORD L. L. , HOLLOWAY Kevin S. , DOMJAN Michael. The nature of sexual reinforcement -- J. Exp. Anal. Behav., 60(1):55-66, 1993
CREWS David. The evolutionary antecedents to love -- Psychoneuroendocrinology, 23(8):751-764, 1998
CUEVA-ROLON Raphael , SANSONE Giorgio , BIANCA Ralph , GOMEZ Lisbeth E. , BEYER Carlos , WHIPPLE Beverly , KOMISARUK Barry R. : Vagotomy blocks responses to vaginocervical stimulation after genitospinal neurectomy in rats ? Physiology and behavior, 60/1:19-24, 1996
CURRIER Richard L. : Juvenile sexuality in global perspective ? in CONSTANTINE Larry L. , MARTINSON Floyd M. , (Ed) : Children and sex : new findings, new perspectives ? Little, Brown and Co, Boston, 1981
CUTLER W. B. , FRIEDMANN E. , MCCOY N. L. Pheromonal influences on sociosexual behavior in men -- Archives of Sexual Behavior, 27(1):1-13, 1998
CYRULNIK Boris : Inhibition et interdiction de l'inceste : approche éthologique ? Psychiatries, 64:35-41, 1985
DALTON P. , DOOLITTLE N. , BRESLIN P. A. Gender-specific induction of enhanced sensitivity to odors -- Nature Neuroscience, 5(3):199-200, 2002
DAMASIO Antonio R. : L'erreur de Descartes : la raison des émotions · Odile Jacob 1995
DAMASIO Antonio R. : Spinoza avait raison : joie et tristesse, le cerveau des émotions · Odile Jacob 2003
DANTZER Robert : Les émotions · PUF QSJ, 1988
DARNELL Jim , LODISH Harvey , BALTIMORE David : Biologie moléculaire de la cellule · De Boeck Université, 2e édition, 1993
DAVIS Michael : The role of the amygdala in fear and anxiety · Annual reviews in neuroscience, 15:353-375, 1992
De AJURIAGUERRA J. Manuel de psychiatrie de l’enfant. Masson, 1970
DE ARAUJO Ivan E. T. , ROLLS Edmund T. , KRINGELBACH Morten L. , MCGLONE Francis , PHILLIPS Nicola. Taste-olfactory convergence, and the representation of the pleasantness of flavour, in the human brain -- Eur. J. Neurosci., 18(7):2059-2068, 2003
DE BONIS Monique : Connaître les émotions humaines · Mardaga 1996
DE RICQLÈS Armand. Théorie synthétique de l'évolution -- Encyclopedia Universalis v. 11. 0, 2005
DE WAAL Frans : De la réconciliation chez les primates ? Flammarion 1992
DE WAAL Frans : L'activité sexuelle pacificatrice des bonobos ? Pour la science, n° spécial, 1996
DE WAAL Frans B. M. Sociosexual behavior used for tension regulation in all age and sex combinations among Bonobos -- Springer-Ferlag, 1990
DEFOLIART Gene. Insects as human food -- Crop Protection, 11(5):395-399, 1992
DEL CERRO M. Cruz R. Role of the vomeronasal input in maternal behavior -- Psychoneuro-endocrinology, 23(8):905-926, 1998
DELACOUR Jean : Science et conscience -- Pour la science, 302:26-32, 2002
DELATTRE Pierre : Interdisciplinaires (recherches) · Encyclopaedia Universalis, (version CD-ROM 5.0, 1999), vol. 12, 1995
DENTON Michael : Evolution : une théorie en crise · Flammarion 1992 (Première publication 1985)
DEPUTTE Bertrand : L'évitement de l'inceste chez les primates ? La recherche 193/18:1332-1342, 1987
DEROCHE-GAMONET V. , BELIN D. , PIAZZA P. V. Evidence for addiction-like behavior in the rat -- Science, 305(5686):1014-1017, 2004
DESCOINS Charles : Phéromones ? Encyclopaedia Universalis, version CD-ROM 6.0 (2000)
DEUTSCH J. , LARSSON K. Model-oriented sexual behavior in surrogate-reared rhesus monkeys -- Brain, Behavior and Evolution, 9(3):157-164, 1974
DEVINSKY O. , MORRELL M. J. , VOGT B. A. Contributions of anterior cingulate cortex to behaviour -- Brain, 118 ( Pt 1):279-306, 1995
DIAMOND Lisa M. What does sexual orientation orient ? A biobehavioral model distinguishing romantic love and sexual desire -- Psychological Review, 110(1):173-192, 2003
DIAMOND M. Sexual behavior in pre contact Hawai'i : a sexological ethnography -- Re vista Española del Pacifico, 16:37-58, 2004
DIAMOND M. , SIGMUNDSON H. K. Sex reassignment at birth. Long-term review and clinical implications -- Arch. Pediatr. Adolesc. Med., 151(3):298-304, 1997
DIAMOND Milton. Bisexuality: A Biological Perspective. in: Erwin J. Haeberle and Rolf Gindorf, eds.: Bisexualities - The ideology and practice of sexual contact with both men and women, New York, Continuum, 1998, pp. 53-80
DODGE Brian , REECE Michael , COLE Sara L. , SANDFORT Theo G. M. Sexual compulsivity among heterosexual college students -- The Journal of Sex Research, 41(4):343-350, 2004
DOMINGUEZ J. M. , HULL E. M. Dopamine, the medial preoptic area, and male sexual behavior -- Physiology & Behavior, 86(3):356-368, 2005
DOMJAN M. , LYONS R. , NORTH N. C. , BRUELL J. Sexual Pavlovian conditioned approach behavior in male Japanese quail (Coturnix coturnix japonica) -- J. Comp Psychol., 100(4):413-421, 1986
DORON Roland , PAROT Françoise (Eds). Dictionnaire de psychologie · PUF 2003
DOUGLAS L. A. , VARLINSKAYA E. I. , SPEAR L. P. Rewarding properties of social interactions in adolescent and adult male and female rats: impact of social versus isolate housing of subjects and partners -- Dev. Psychobiol., 45(3):153-162, 2004
DU J. , HULL E. M. Effects of testosterone on neuronal nitric oxide synthase and tyrosine hydroxylase -- Brain Research, 836(1-2):90-98, 1999
DULAC C. , TORELLO A. T. Molecular detection of pheromone signals in mammals: from genes to behaviour -- Nat. Rev. Neurosci., 4(7):551-562, 2003
DUPANLOUP Isabelle , PEREIRA Luisa , BERTORELLE Giorgio. , CALAFELL F. , PRATA M. J. , AMORIM A. , BARBUJANI G. A recent shift from polygyny to monogamy in humans is suggested by the analysis of worldwide Y-chromosome diversity -- Journal of Molecular Evolution, 57(1):85-97, 2003
DUPONT R. L. Addiction: a new paradigm -- Bull. Menninger Clin., 62(2):231-242, 1998
EDELMAN Gerald M. : Biologie de la conscience -- Odile Jacob 1992
EDGERTON Robert B. Sick societies. Challenging the myth of primitive harmony -- Macmillan, 1992
EKE N. Fracture of the penis -- British Journal of Surgery, 89(5):555-565, 2002
ERICKSON M. T. Rethinking Oedipus: an evolutionary perspective of incest avoidance -- The American journal of psychiatry, 150(3):411-416, 1993
EVANS Terry. Bisexuality : negotiating lives between two cultures -- Journal of bisexuality, 3(2):91-108, 2003
EVERITT Barry J. : Sexual motivation : a neural and behavioral analysis of the mechanisms underlying appetitive and copulatory response of male rats ? Neuroscience and biobehavioral reviews, 14:217-232, 1990
FERRIS C. F. , KULKARNI P. , SULLIVAN J. M., Jr. , HARDER J. A. , MESSENGER T. L. , FEBO M. Pup suckling is more rewarding than cocaine: evidence from functional magnetic resonance imaging and three-dimensional computational analysis -- The Journal of Neuroscience, 25(1):149-156, 2005
FERVEUR Jean-François , SAVARI Fabrice , O'KANE Cahir J. , SUREAU Gilles , GREENSPAN Ralph , JALLON Jean-Marc : Genetic feminization of pheromones and its behavioral consequences in Drosophila males ? Science, 276:1555-1558, 1997
FEYERABEND Paul : Contre la méthode : esquisse d'une théorie anarchiste de la connaissance · Seuil, Paris, 1979
FIELD Tiffany. Massage therapy for infants and children. Journal of developmental and behavioral pediatrics, 16/2:105-111, 1995
FILSINGER EE, BRAUN JJ, MONTE WC. An examination of the effects of putative pheromones on human judgments · Ethol Sociobiol 1985; 6:227–36
FIRESTEIN Stuart : How the olfactory system makes sense of scents · Nature, 413:211-218, 2001
FISHER Helen , ARON Arthur , MASHEK Debra , LI Haifang , BROWN Lucy L. Defining the brain systems of lust, romantic attraction, and attachment -- Archives of Sexual Behavior, 31(5):413-419, 2002
FISHER Helen , ARON Arthur , MASHEK Debra , LI Haifang , STRONG Greg , BROWN Lucy L. The Neural Mechanisms of Mate Choice: A Hypothesis -- Neuroendocrinology Letters, 23 Suppl 4:92-97, 2002
FISKE Brian. The sexual brain -- Nature Neuroscience, 7(10):1029, 2004
FLANDRIN Jean-louis : La vie sexuelle des gens mariés dans l'ancienne société ? Communications, 35:102-115, 1982
FOIDART A. , LEGROS J.J. , BALTHAZART J. : Les phéromones humaines : vestige animal ou réalité non reconnue ? Revue médicale de Liège, 49/12:662-680, 1994
FORD Clellan S. , BEACH Frank A. : Patterns of sexual behavior ? Methuen & Co, London, 1965
FOUCAULT Michel : Le combat de la chasteté ? Communications, 35:15-25, 1982
FOX Robin : Les conditions de l'évolution sexuelle ? Communications, 35:2-14, 1982
FRANCIS S. , ROLLS Edmund T. , BOWTELL R. , MCGLONE F. , O'DOHERTY J. , BROWNING A. , CLARE S. , SMITH E. The representation of pleasant touch in the brain and its relationship with taste and olfactory areas -- Neuroreport, 10(3):453-459, 1999
FRAYSER S. G. Defining normal childhood sexuality : an anthropological approach -- Annual Review of Sex Research, 5:173-217, 1994
FREUD Sigmund. Trois Essais sur la théorie de la sexualité · Gallimard, 1962
FRIEDMAN H. L. Changing patterns of adolescent sexual behavior : consequences for health and development -- Journal of Adolescent Health, 13(5):345-350, 1992
FRIEDRICH William N. , GRAMBSCH Patricia. Child sexual behavior inventory : normative and clinical comparisons -- Psychological Assessment, 4(3):303-311, 1992
FRITH Chris D. , FRITH Uta. Interacting minds--a biological basis -- Science, 286(5445):1692-1695, 1999
FROHLICH Jonathan , OGAWA Sonoko , MORGAN Maria , BURTON Leslie , PFAFF Donald. Hormones, genes and the structure of sexual arousal -- Behavioural Brain Research, 105(1):5-27, 1999
GADPAILLE Warren J. : The delay of normal psychosexual development ? in CONSTANTINE Larry L. , MARTINSON Floyd M. , (Ed) : Children and sex : new findings, new perspectives ? Little, Brown and Co, Boston, 1981
GALLO A. & COQUERY J.-M. p.388 in BLOCH Henriette , CHEMAMA Roland , GALLO Alain , LECONTE Pierre , LE NY Jean-François , POSTEL Jacques , MOSCOVICI Serge , REUCHLIN Maurice , VURPILLOT Eliane , (Ed) : Grand dictionnaire de la psychologie -- Larousse 1994
GANDELMAN R. : Gonadal hormone and sensory function -- Neurocience and biobehavioral reviews, 7:1-18, 1983
GARIEPY Jean-Louis , LEWIS Mark H. , CAIRNS Robert B. : Genes, neurobiology, and aggression : time frames and functions of social behaviors in adaptation -- in STOFF David M. , CAIRNS Robert B. : Aggression and violence : genetic, neurobiological, and biosocial perspectives -- Lawrence Erlbaum Associates, New Jersey, 1996
GAY Judith : "Mummies and babies" and friends and lovers in Lesotho ? in BLACKWOOD Evelyne (Ed) : The many faces of homosexuality : anthropological approaches to homosexual behavior ? Harrington Park Press, New York, 1986
GAZZANIGA Michael S. (Ed) : The cognitive neurosciences · Bradford book, Cambridge, 1995
GAZZANIGA Michael S. , IVRY Richard B. , MANGUN George R. : Cognitive neuroscience : the biology of the mind -- Norton & Company, 2nd edition, 2002
GÉNERMONT Jean , BOCQUET Charles. Adaptation biologique -- Encyclopedia Universalis v. 11. 0, 2005
GEORGE Mark S. , KETTER Terence A. , PAREKH Priti I. , HERSCOVITCH Peter , POST Robert M. Gender differences in regional cerebral blood flow during transient self-induced sadness or happiness -- Biological Psychiatry, 40(9):859-871, 1996
GEORGE Mark S. , KETTER Terence A. , PAREKH Priti I. , HORWITZ B. , HERSCOVITCH Peter , POST Robert M. Brain activity during transient sadness and happiness in healthy women -- The American journal of psychiatry, 152(3):341-351, 1995
GERALL H. D. , WARD I. L. , GERALL A. A. Disruption of the male rat's sexual behaviour induced by social isolation -- Anim Behav., 15(1):54-58, 1967
GIL Fernando : Preuve (épistémologie) · Encyclopaedia Universalis, (version CD-ROM 5.0, 1999), vol. 18, 1995
GILBERT A.N. , YAMAZAKI K. , BEAUCHAMP G.K. , THOMAS L. : Olfactory discrimination of mouse strains (mus musculus) and major histocompatibility types by humans (homo sapiens) ? Journal of comparative psychology, 100/3, 262-265, 1986
GILBERT Scott F. : Biologie du développement · De Boeck Université 1996
GIORGI G. , SICCARDI M. Ultrasonographic observation of a female fetus' sexual behavior in utero -- Am. J. Obstet. Gynecol., 175(3 Pt 1):753, 1996
GIULIANO F. , RAMPIN O. Neural control of erection -- Physiology & Behavior, 83(2):189-201, 2004
GIULIANO F. , TOSTAIN J. , ROSSI D. Testosterone and male sexuality: basic research and clinical data -- Prog. Urol., 14(5):783-790, 2004
GIULIANO François , RAMPIN Olivier , BERNABE Jacques , JARDIN Alain , BENOIT Gérard : Approche expérimentale de l'érection réflexe chez le rat : modélisation et neuroanatomie fonctionnelle des voies nerveuses impliquées ? Progrès en urologie, 6:81-86, 1996
GOGATE M. G. , BRID S. V. , WINGKAR K. C. , KANTAK N. M. Septal regulation of male sexual behavior in rats -- Physiology & Behavior, 57(6):1205-1207, 1995
GOLDFOOT D. A. Rearing conditions which support or inhibit later sexual potential of laboratory-born rhesus monkeys: hypothesis and diagnostic behaviors -- Lab Anim Sci., 27(4):548-556, 1977
GOLDFOOT David A. Olfaction, sexual behavior, and the pheromone hypothesis in rhesus monkeys A critique -- American Zoologist, 21(1):153-164, 1981
GOODMAN A. Addiction: definition and implications -- Br. J. Addict., 85(11):1403-1408, 1990
GOODMAN A. Diagnosis and treatment of sexual addiction -- Journal of Sex and Marital Therapy, 19(3):225-251, 1993
GOODMAN A. Sexual addiction: designation and treatment -- Journal of Sex and Marital Therapy, 18(4):303-314, 1992
GOULD Stephen Jay. Quand les poules auront des dents. Réflexions sur l'histoire naturelle -- Fayard, 1991
GOWER D.B. , RUPARELIA B.A. : Olfaction in humans with special reference to odorous 16-androstenes : their occurence, perception and possible social, psychological, and sexual impact ? Journal of endocrinology, 137:167-187, 1993
GRAMBSCH P. , GRAMBSCH Patricia , BROUGHTON Daniel , KUIPER James , BEILKE Robert L. Normative sexual behavior in children -- Pediatrics, 88(3):456-464, 1991
GRAMMER Karl. Human courtship behavior : biological basis and cognitive processing -- in RASA A. E. , VOGEL C. , VOLAND E. (Eds). The sociobiology of sexual and reproductive strategies -- Chapmann and Hall, (9):147-169, 1989
GRASSÉ Pierre-Paul , PETIT Claudine , FORTERRE Patrick. Evolution -- Encyclopedia Universalis v. 11. 0, 2005
GRAY Alison , PITHERS William D. , BUSCONI Aida , HOUCHENS Paul. Developmental and etiological characteristics of children with sexual behavior problems : treatment implications -- Child Abuse and Neglect, 23(6):601-621, 1999
GREENE J. D. , NYSTROM L. E. , ENGELL A. D. , DARLEY J. M. , COHEN J. D. The neural bases of cognitive conflict and control in moral judgment -- Neuron, 44(2):389-400, 2004
GRIMOULT Cédric. Mutationnisme -- Encyclopedia Universalis v. 11. 0, 2005
GRUENDEL A. D. , ARNOLD W. J. Effects of early social deprivation on reproductive behavior of male rats -- J. Comp Physiol Psychol., 67(1):123-128, 1969
GUÉGUEN Nicolas. Un parfum de séduction -- Cerveau & Psycho, (17):30-32, 2006
GUILLEBAUD Jean-Claude : La tyrannie du plaisir ? Seuil 1998
GUNDERSEN Bjorn Helge , MELAS Per Steinar , SKAR Jens E. : Sexual behavior of preschool children : teachers' observations ? in CONSTANTINE Larry L. , MARTINSON Floyd M. , (Ed) : Children and sex : new findings, new perspectives ? Little, Brown and Co, Boston, 1981
GUYTON Arthur C. : Anatomie et physiologie du système nerveux · Decarie Vigot 1989
HAIG David. The inexorable rise of gender and the decline of sex : social change in academic titles, 1945-2001 -- Archives of Sexual Behavior, 33(2):87-96, 2004
HALATA Z. , MUNGER B.L. : The neuroanatomical basis for the protopathic sensibility of the human glans penis ? Brain research, 371/2:205-230, 1986
HALPERN Carolyn T. , JOYNER Kara , UDRY J. Richard , SUCHINDRAN Chirayath. Smart teens don't have sex (or kiss much either) -- Journal of Adolescent Health, 26(3):213-225, 2000
HAMMOND C. , TRITSCH D. : Neurobiologie cellulaire · Doin 1990
HARD E. , LARSSON K. Climbing behavior patterns in prepubertal rats. Effects of hormones, rearing conditions, and sensory cues -- Brain, Behavior and Evolution, 4(2):151-161, 1971
HASHIMOTO Chie. Context and development of sexual behavior of wild bonobos (pan paniscus) at Wamba, Zaire -- International Journal of Primatology, 18(1):1-21, 1997
HATFIELD E. , SPRECHER S. Measuring passionate love in intimate relationships -- J. Adolesc., 9(4):383-410, 1986
HEATH Robert G. : Pleasure and brain activity in man ? The journal of nervous and mental disease, 154/1:3-18, 1972
HEDRICKS Cynthia A. Female sexual activity across the human menstrual cycle -- Annual Review of Sex Research, 5:122-172, 1994
HENDRICKS Shelton E. : Role of estrogen and progestins in the development of female sexual behavior potential ? in GERAL Arnold A. , MOLTZ Howard , WARD Ingeborg L. (Ed) : Sexual differentiation, vol 11, Handbook of behavioral neurobiology ? Plenum Press, NY, 1992
HENRY James P. , WANG Sheila. Effects of early stress on adult affiliative behavior -- Psychoneuroendocrinology, 23(8):863-875, 1998
HENRY Jules , HENRY Zunia. Doll play of Pilaga indian children -- First Vintage Books Edition, 1974
HÉRAULT Jeanny, JUTTEN Christian. Réseaux neuronaux et traitement du signal. Hermes, 1994
HERDT Gilbert , MCCLINTOCK Martha K. The magical age of 10 -- Archives of Sexual Behavior, 29(6):587-606, 2000
HERDT Gilbert , STOLLER Robert J. Intimate communications: erotics and the study of culture -- Columbia University Press, NY, 1990
HERMAN-JEGLINSKA Anna , GRABOWSKA Anna , DULKO Stanislaw. Masculinity, femininity, and transsexualism -- Archives of Sexual Behavior, 31(6):527-534, 2002
HILL Craig A. , PRESTON Leslie K. Individual differences in the experience of sexual motivation : theory and measurement of dispositional sexual motives -- The Journal of Sex Research, 33(1):27-45, 1996
HINES T. M. The G-spot: a modern gynecologic myth -- Am. J. Obstet. Gynecol., 185(2):359-362, 2001
HOANG N. M. , SMADJA A. , HERVE DE SIGALONY J. P. The reality and usefulness of Halban's fascia -- J. Gynecol. Obstet. Biol. Reprod. (Paris), 20(1):51-59, 1991
HOLE G. J. , EINON D. F. , PLOTKIN H. C. The role of social experience in the development of sexual competence in rattus norvegicus -- Behavioural Process, 12:187-202, 1986
HOLLAND Peter C. , GALLAGHER Michela. Amygdala-frontal interactions and reward expectancy -- Current Opinion in Neurobiology, 14(2):148-155, 2004
HOLLEY André : L'information olfactive et son traitement neuronal ? Revue internationale de psychopathologie, 22:305-337, 1996
HOLLOWAY Kevin S. , DOMJAN Michael. Sexual approach conditioning : tests of unconditioned stimulus devaluation using hormone manipulations -- J. Exp. Psychol. Anim Behav. Process, 19(1):47-55, 1993
HOLLOWAY Kevin S. , DOMJAN Michael. Sexual approach conditioning : unconditioned stimulus factors -- J. Exp. Psychol. Anim Behav. Process, 19(1):38-46, 1993
HOLSTEGE G. , GEORGIADIS J. R. , PAANS A. M. , MEINERS L. C. , VAN DER GRAAF F. H. , REINDERS A. A. Brain activation during human male ejaculation -- The Journal of Neuroscience, 23(27):9185-9193, 2003
HOLSTEGE G. , BANDLER R. , SAPER C.B. : The emotional motor system · Elsevier 1996
HOROWITZ Stephen M. , WEIS David L. , LAFLIN Molly T. Differences between sexual orientation behavior groups and social background, quality of life, and health behaviors - Statistical Data Included -- The Journal of Sex Research, 38(3):205-218, 2001
HOUDART Raymond. L'affectivité dans le système nerveux. L'Encéphale, XXX:236-244, 2004
HULL Elaine M. , LORRAIN Daniel S. , DU Jianfang , MATUSZEWICH Leslie , LUMLEY L. A. , PUTNAM S. K. , MOSES J. Hormone-neurotransmitter interactions in the control of sexual behavior -- Behavioural Brain Research, 105(1):105-116, 1999
INSEL Thomas R. Is social attachment an addictive disorder? -- Physiology & Behavior, 79(3):351-357, 2003
INSEL Thomas R. , GELHARD R. , SHAPIRO L. E. The comparative distribution of forebrain receptors for neurohypophyseal peptides in monogamous and polygamous mice -- Neuroscience, 43(2-3):623-630, 1991
INSEL Thomas R. , GINGRICH B. S. , YOUNG L. J. Oxytocin : who needs it? -- Progress in Brain Research, 133:59-66, 2001
INSEL Thomas R. , PRESTON Stephanie , WINSLOW James T. Mating in the monogamous male: behavioral consequences -- Physiology & Behavior, 57(4):615-627, 1995
INSEL Thomas R. , SHAPIRO Lawrence E. Oxytocin receptor distribution reflects social organization in monogamous and polygamous voles -- Proceedings of the National Academy of Sciences of the United States of America, 89(13):5981-5985, 1992
INSEL Thomas R. , WINSLOW James T. Central administration of oxytocin modulates the infant rat's response to social isolation -- Eur. J. Pharmacol., 203(1):149-152, 1991
INSEL Thomas R. , YOUNG Larry , WANG Zuoxin : Molecular aspects of monogamy ? Ann N Y Acad Sci, 807:302-316, 1997
INSEL Thomas R. Oxytocin - a neuropeptide for affiliation : evidence from behavioral, receptor autoradiographic, and comparative studies. Psychoneuroendocrinology, 17/1:3-35, 1992
JACOB François. Evolution and tinkering -- Science, 196:1161-1166, 1977
JACOB François. Le jeu des possibles -- Fayard, 1981
JAKOBOVITS A. A. Fetal penile erection -- Ultrasound in Obstetrics and Gynecology, 18(4):405, 2001
JANKOWIAK W. R. , FISCHER E. F. A cross-cultural perspective on romantic love -- Ethnology, 31:149, 1992
JANUS Samuel S. , BESS Barbara E. : Latency : fact or fiction ? ? in CONSTANTINE Larry L. , MARTINSON Floyd M. , (Ed) : Children and sex : new findings, new perspectives ? Little, Brown and Co, Boston, 1981
JEANNEROD Marc : Traitement conscient et inconscient de l'information perceptive -- Revue internationale de psychopathologie, I:13-34, 1990
JENKINS W. J. , BECKER J. B. Role of the striatum and nucleus accumbens in paced copulatory behavior in the female rat -- Behavioural Brain Research, 121(1-2):119-128, 2001
JOHNSON P. , MAXWELL D. Fetal penile length -- Ultrasound in Obstetrics and Gynecology, 15(4):308-310, 2000
JOHNSON Scott P. , AMSO Dima , SLEMMER Jonathan A. Development of object concepts in infancy: Evidence for early learning in an eye-tracking paradigm -- Proceedings of the National Academy of Sciences of the United States of America, 100(18):10568-10573, 2003
JOHNSTON Charley M. , DEISHER Robert W. : Contemporary communal child rearing : a first analysis · Pediatrics, 52/3:319-326, 1973
JONES Edward G. Cortical and subcortical contributions to activity-dependent plasticity in primate somatosensory cortex -- Annu. Rev. Neurosci., 23:1-37, 2000
JORDAN Bertrand : Histoire d'X : anatomie de deux dérives : du gène de l'homosexualité à celui de la criminalité ? La recherche, 311:84-88, 1998
KAHLE W. , LEONHARDT H. , PLATZER W. : Anatomie : système nerveux · Flammarion-médecine-sciences 1992
KALICHMAN Seth C. , ROMPA David. Sexual sensation seeking and Sexual Compulsivity Scales: reliability, validity, and predicting HIV risk behavior -- J. Pers. Assess., 65(3):586-601, 1995
KANDEL Eric R. , SCHWARTZ James H. : Essentials of neural sciences and behavior · Appleton et Lange, Norwalk Connecticut 1995
KARAMA Sherif , LECOURS Andre R. , LEROUX Jean-Maxime , BOURGOUIN Pierre , BEAUDOIN Gilles , JOUBERT Sven , BEAUREGARD Mario. Areas of brain activation in males and females during viewing of erotic film excerpts -- Human Brain Mapping, 16(1):1-13, 2002
KARLI Pierre : L'homme agressif -- Odile Jacob 1987
KELLEY Ann E. , BERRIDGE Kent C. The neuroscience of natural rewards: relevance to addictive drugs -- The Journal of Neuroscience, 22(9):3306-3311, 2002
KELLY D. R. When is a butterfly like an elephant? -- Chem. Biol., 3(8):595-602, 1996
KENDRICK K. M. , HINTON M. R. , ATKINS K. , HAUPT M. A. , SKINNER J. D. Mothers determine sexual preferences -- Nature, 395(6699):229-230, 1998
KEVERNE E. B. Importance of olfactory and vomeronasal systems for male sexual function -- Physiology & Behavior, 83(2):177-187, 2004
KING Brian Edward , ALLGEIER Elizabeth Rice. The Sexual Desire Inventory as a measure of sexual motivation in college students -- Psychological Reports, 86(1):347-350, 2000
KINSEY Alfred C., POMEROY Wardell B., MARTIN Clyde E. Le comportement sexuel de l'homme. Rayonnement de la pensée, 1948
KLEIN Fritz : The bisexual option ? Harrington Park Press, 2nd edition, 1993
KNOBIL Ernest , NEILL Jimmy D. : The physiology of reproduction ? Raven Press, 2nd edition, 1994
KOESTER Susan E. , O'LEARY Dennis D.M. : Development of projection neurons of the mammalian cerebral cortex · in VAN PELT J. , CORNER M.A. , UYLINGS H.B.M. , LOPES DA SILVA F.H. , (Ed) : The self-organizing brain : from growth cones to functional networks · Elsevier 1994
KOHL James V. , ATZMUELLER Michaela , FINK Bernhard , GRAMMER Karl. Human pheromones : integrating neuroendocrinology and ethology -- Neuroendocrinology Letters, 22(5):309-321, 2001
KOLLACK-WALKER S. , NEWMAN S. W. Mating and agonistic behavior produce different patterns of Fos immunolabeling in the male Syrian hamster brain -- Neuroscience, 66(3):721-736, 1995
KOMISARUK Barry R. , BIANCA Ralph , SANSONE Giorgio , GOMEZ Lisbeth E. , CUEVA-ROLON Raphael , BEYER Carlos , WHIPPLE Beverly : Brain-mediated responses to vaginocervical stimulation in spinal cord-transected rats : role of the vagus nerves ? Brain research, 708:128-134, 1996
KOOB G. F. , LE MOAL M. Drug addiction, dysregulation of reward, and allostasis -- Neuropsychopharmacology, 24(2):97-129, 2001
KOVACS Gabor L. , SARNYAI Zoltan , SZABO Gyula. Oxytocin and addiction : a review -- Psychoneuroendocrinology, 23(8):945-962, 1998
KOVACS Gyula , GULYAS Balazs , SAVIC Ivanka , PERRETT D. I. , CORNWELL R. E. , LITTLE A. C. , JONES B. C. , BURT D. M. , GAL V. , VIDNYANSZKY Z. Smelling human sex hormone-like compounds affects face gender judgment of men -- Neuroreport, 15(8):1275-1277, 2004
KOYANAGI T. , HORIMOTO N. , NAKANO H. REM sleep determined using in utero penile tumescence in the human fetus at term -- Biology of the Neonate, 60 Suppl 1:30-35, 1991
KRAFFT-EBING Richard. Psychopathia sexualis -- Agora, 1999
KRINGELBACH Morten L. Food for thought: hedonic experience beyond homeostasis in the human brain -- Neuroscience, 126(4):807-819, 2004
KRINGELBACH Morten L. , O'DOHERTY J. , ROLLS Edmund T. , ANDREWS C. Activation of the human orbitofrontal cortex to a liquid food stimulus is correlated with its subjective pleasantness -- Cerebral Cortex, 13(10):1064-1071, 2003
KRIVACSKA J. J. Child sexual abuse prevention programs : The prevention of childhood sexuality ? -- Journal of Child Sexual Abuse, 1(4):83-112, 1992
KRUGER Tillmann H. , HAAKE Philip , HARTMANN Uwe , SCHEDLOWSKI Manfred , EXTON Michael S. Orgasm-induced prolactin secretion : feedback control of sexual drive ? -- Neuroscience and Biobehavioral Reviews, 26(1):31-44, 2002
KUHN Thomas S. : La structure des révolutions scientifiques · Champs Flammarion 1983
LABARDERA J. D. , MARTIN J. E. , DOZIER J. E. Child's psychiatrist's view of father-daughter incest -- Child Abuse and Neglect, 4:147-151, 1980
LABAT J. J. , MAUDUYT DE LA GRÈVE I. Les troubles génito-sexuels du spina-bifida paralytique -- in COSTA Pierre , LOPEZ S. , PÉLISSIER J. (Eds). Sexualité, fertilité et handicap -- :161-169, 1996
LADRIÈRE Jean : Sciences et discours rationnel · Encyclopaedia Universalis, (version CD-ROM 5.0, 1999), vol. 20, 1995
LAGET Paul. Réflexes · Encyclopaedia Universalis, version CD-ROM 11.0, 2005
LANDOLT Monica A. , BARTHOLOMEW Kim , SAFFREY Colleen , ORAM Doug , PERLMAN Daniel. Gender nonconformity, childhood rejection, and adult attachment : a study of gay men -- Archives of Sexual Behavior, 33(2):117-128, 2004
LANE R. D. , REIMAN Eric M. , AXELROD B. , YUN L. S. , HOLMES A. , SCHWARTZ G. E. Neural correlates of levels of emotional awareness. Evidence of an interaction between emotion and attention in the anterior cingulate cortex -- Journal of Cognitive Neuroscience, 10(4):525-535, 1998
LANE Richard D. , REIMAN Eric M. , AHERN G. L. , SCHWARTZ G. E. , DAVIDSON R. J. Neuroanatomical correlates of happiness, sadness, and disgust -- The American journal of psychiatry, 154(7):926-933, 1997
LANE Richard D. , REIMAN Eric M. , BRADLEY Margaret M. , LANG P. J. , AHERN G. L. , DAVIDSON R. J. , SCHWARTZ G. E. Neuroanatomical correlates of pleasant and unpleasant emotion -- Neuropsychologia, 35(11):1437-1444, 1997
LANGANEY André , NADOT Robert : Génétique, parenté et prohibition de l'inceste ? in DUCROS, Albert ; PANOFF, Michel : la frontière des sexes ? PUF 1995
LANGFELDT Thore : Childhood masturbation : individual and social organization ? in CONSTANTINE Larry L. , MARTINSON Floyd M. , (Ed) : Children and sex : new findings, new perspectives ? Little, Brown and Co, Boston, 1981
LANGFELDT Thore : Progress in sexual development ? in CONSTANTINE Larry L. , MARTINSON Floyd M. , (Ed) : Children and sex : new findings, new perspectives ? Little, Brown and Co, Boston, 1981
LARGEAULT Jean : Description et explication · Encyclopaedia Universalis, version CD-ROM 5.0, 1999
LARGEAULT Jean : Méthode · Encyclopaedia Universalis, (version CD-ROM 5.0, 1999), vol. 15, 1995
LAUMANN E. O. , PAIK A. , ROSEN R. C. Sexual dysfunction in the United States: prevalence and predictors -- JAMA, the Journal of the American Medical Association, 281(6):537-544, 1999
LAUREYS Steven , FAYMONVILLE Marie-Élisabeth , MAQUET Pierre : Quelle conscience durant le coma -- Pour la science, 302:122-128, 2002
LE DOUX J.E. : In search of an emotional system in the brain. in GAZZANIGA Michael S. (Ed) : The cognitive neurosciences · Bradford book, Cambridge, 1995
LE GALL A. , MULLET E. , RIVIERE Shafighi S. Age, religious beliefs, and sexual attitudes -- The Journal of Sex Research, 39(3):207-216, 2002
LEAK Rehana K. , MOORE robert Y. : Identification of retinal ganglion cells projecting to the lateral hypothalamic area of the rat · Brain research, 770:105-114, 1997
LECANUET J.P., GRANIER-DEFERRE C., SCHAAL B. Continuité sensorielle transnatale. in POUTHAS Viviane, JOUEN François. Les comportements du bébé : expression de son savoir ? Mardaga, Liège, 1993 p34
LECKMAN J. F. , FELDMAN R. , SWAIN J. E. , EICHER V. , THOMPSON N. , MAYES L. C. Primary parental preoccupation: circuits, genes, and the crucial role of the environment -- Journal of Neural Transmission, 111(7):753-771, 2004
LECKMAN J. F. , MAYES L. C. Preoccupations and behaviors associated with romantic and parental love. Perspectives on the origin of obsessive-compulsive disorder -- Child and Adolescent Psychiatric Clinics of North America, 8(3):635-665, 1999
LECKMAN J. F. , MAYES L. C. , FELDMAN R. , EVANS D. W. , KING R. A. , COHEN D. J. Early parental preoccupations and behaviors and their possible relationship to the symptoms of obsessive-compulsive disorder -- Acta psychiatrica scandinavica. Supplementum, 396:1-26, 1999
LEDOUX J. E. Emotion circuits in the brain -- Annu. Rev. Neurosci., 23:155-184, 2000
LEIDERMAN Herbert P., BABU Béatrice, KAGIA James, KRAEMER Helena C., LEIDERMAN Gloria F. African infant precocity and some social influences during the first year. Nature, 242:247-249, 1973
LEITENBERG Harold , GREENWALD E. , TARRAN M. J. The relation between sexual activity among children during preadolescence and/or early adolescence and sexual behavior and adjustement in young adulthood -- Archives of Sexual Behavior, 18(4):299-313, 1989
LEITENBERG Harold , SALTZMAN Heidi. A statewide survey of age at first intercourse for adolescent females and age of their male partners: relation to other risk behaviors and statutory rape implications -- Archives of Sexual Behavior, 29(3):203-215, 2000
LEITENBERG Harold , SALTZMAN Heidi. College women who had sexual intercourse when they were underage minors (13-15): age of their male partners, relation to current adjustment, and statutory rape implications -- Sex Abuse, 15(2):135-147, 2003
LENCK L. C. , VANNEUVILLE G. , MONNET J. P. , HARMAND Y. Urethral sphincter (G point). Anatomo-clinical correlations -- Rev. Fr. Gynecol. Obstet., 87(2):65-69, 1992
LENDERKING W. R. , WOLD C. , MAYER K. H. , GOLDSTEIN R. , LOSINA E. , SEAGE G. R., III. Childhood sexual abuse among homosexual men. Prevalence and association with unsafe sex -- J. Gen. Intern. Med., 12(4):250-253, 1997
LEVAY S. A difference in hypothalamic structure between heterosexual and homosexual men -- Science, 253(5023):1034-1037, 1991
LEVER Maurice : Les bûchers de Sodome ? Fayard 10/18, 1996
LEVESQUE Johanne , JOANETTE Y. , MENSOUR Boualem , BEAUDOIN Gilles , LEROUX Jean-Maxime , BOURGOUIN Pierre , BEAUREGARD Mario. Neural correlates of sad feelings in healthy girls -- Neuroscience, 121(3):545-551, 2003
LEVESQUE Johanne , JOANETTE Y. , MENSOUR Boualem , BEAUDOIN Gilles , LEROUX Jean-Maxime , BOURGOUIN Pierre , BEAUREGARD Mario. Neural basis of emotional self-regulation in childhood -- Neuroscience, 129(2):361-369, 2004
LEVIN Roy J. , VAN BERLO Willy. Sexual arousal and orgasm in subjects who experience forced or non-consensual sexual stimulation - a review -- Journal of Clinical Forensic Medicine, 11(2):82-88, 2004
LEVINE S. B. An essay on the nature of sexual desire -- Journal of Sex and Marital Therapy, 10(2):83-96, 1984
LIBERLES S. D. , BUCK L. B. A second class of chemosensory receptors in the olfactory epithelium -- Nature, 442(7103):645-650, 2006
LIMAN Emily R. : Pheromone transduction in the vomeronasal organ ? Current opinion in neurobiology, 6:487-493, 1996
LIU Yiang-cheng , SALAMONE John D. , SACHS Benjamin D. : Lesions in medial preoptic area and bed nucleus of stria terminalis : differential effects on copulatory behavior and non contact erection in male rats ? The journal of neuroscience, 17/13:5245-5253, 1997
LOONIS Eric , SZTULMAN Henry. [Is the functioning of our brain of an addictive nature?] -- Encephale, 24(1):26-32, 1998
LOVEJOY C. O. The origin of man -- Science, 211(4480):341-350, 1981
LOWRY T. P. How breast feeding arouses woman -- Journal of Sexology, 37:46-49, 2004
MADDOCK R. J. The retrosplenial cortex and emotion: new insights from functional neuroimaging of the human brain -- Trends in Neurosciences, 22(7):310-316, 1999
MAGGIO J. C. , MAGGIO J. H. , WHITNEY G. Experience-based vocalization of male mice to female chemosignals -- Physiology & Behavior, 31(3):269-272, 1983
MAH Kenneth, BINIK Yitzchack M. : The nature of human orgasm : a critical review of major trends ? Clinical Psychology Review, 21/6:823-856, 2001
MAH Kenneth. , BINIK Y. M. Do all orgasms feel alike ? Evaluating a two-dimensional model of the orgasm experience across gender and sexual context -- The Journal of Sex Research, 39(2):104-113, 2002
MAIWORM RE. Influence of androstenone, androstenol, menstrual cycle, and oral contraceptives on the attractivity ratings of female probands · Paper presented at the Ninth Congress of ECRO; 1990
MALINOVSKY L. Sensory nerve formations in the skin and their classification -- Microsc. Res. Tech., 34(4):283-301, 1996
MALINOWSKI Bronislaw : la vie sexuelle des sauvages du nord-ouest de la mélanésie ? Petite bibliothèque Payot 1970
MALLICK H.N. , MANCHANDA S.K. , KUMAR Mohan V. : Sensory modulation of the medial preoptic area neuronal activity by dorsal penile nerve stimulation in rats ? The journal of urology, 151:759-762, 1994
MARAZZITI D. , CASSANO G. B. The neurobiology of attraction -- Journal of Endocrinological Investigation, 26(3 Suppl):58-60, 2003
MARIEB Elaine N. : Anatomie et physiologie humaine · De Boeck Université, Bruxelle, 2e édition, 1993
MARSHALL Leslie B. Infant care and feeding in the South Pacific -- Gordon and Breach, NY, 1985
MARSON Lesley , MAC KENNA Kevin E. : Stimulation of the hypothalamus initiates the urethrogenital reflex in male rats ? Brain research, 638:103-108, 1994
MARSON Lesley , MAC KENNA Kevin E. : The identification of a brainstem site controllign spinal sexual reflexes in male rats ? Brain research, 515:303-308, 1990
MARSON Lesley , PLATT K.B. , MAC KENNA Kevin E. : Central nervous system innervation of the penis as revealed by the transneuronal transport of pseudorabies virus ? Neuroscience, 55/1:263-280, 1993
MARSON Lesley. Central nervous system neurons identified after injection of pseudorabies virus into the rat clitoris -- Neuroscience Letters, 190(1):41-44, 1995
MARTINSON Floyd M. : The sexual life of children ? Bergin & Garvey, 1994
MARTINSON(a) Floyd M. : Childhood and the institutionalization of sexuality ? in CONSTANTINE Larry L. , MARTINSON Floyd M. , (Ed) : Children and sex : new findings, new perspectives ? Little, Brown and Co, Boston, 1981
MARTINSON(b) Floyd M. : Eroticism in infancy and childhood ? in CONSTANTINE Larry L. , MARTINSON Floyd M. , (Ed) : Children and sex : new findings, new perspectives ? Little, Brown and Co, Boston, 1981
MASON William A. , MENDOZA Sally P. Generic aspects of primate attachments : parents, offspring and mates -- Psychoneuroendocrinology, 23(8):765-778, 1998
MASTERS William H. , JOHNSON Virginia E. , KOLODNY Robert C. : Amour et sexualité : mieux vivre sa vie sexuelle dans le monde d'aujourd'hui ? Interéditions, 1987
MASTERS William H. , JOHNSON Virginia E. : Human sexual response · Bantam Books 1980 (première publication 1966)
MAZOYER Bernard , ZAGO L. , MELLET E. , BRICOGNE S. , ETARD O. , HOUDE O. , CRIVELLO F. , JOLIOT M. , PETIT L. , TZOURIO-MAZOYER N. Cortical networks for working memory and executive functions sustain the conscious resting state in man -- Brain Research Bulletin, 54(3):287-298, 2001
MCBRIDE W. J. , MURPHY J. M. , IKEMOTO S. Localization of brain reinforcement mechanisms: intracranial self-administration and intracranial place-conditioning studies -- Behavioural Brain Research, 101(2):129-152, 1999
MCCABE M. P. , CUMMINS R. A. Sexuality and quality of life among young people -- Adolescence, 33(132):761-773, 1998
MCCLELLAN J. , MCCURRY C. , RONNEI M. , ADAMS J. , EISNER A. , STORCK M. Age of onset of sexual abuse : relationship to sexually inappropriate behaviors -- Journal of the American Academy of Child and Adolescent Psychiatry, 35(10):1375-1383, 1996
MCCLINTOCK Martha K. , HERDT Gilbert. Rethinking puberty : The development of sexual attraction -- Current Developments in Psychological Science, 5(6):178-183, 1996
MCCLURE S. M. , LAIBSON D. I. , LOEWENSTEIN G. , COHEN J. D. Separate neural systems value immediate and delayed monetary rewards -- Science, 306(5695):503-507, 2004
MCCLURE Samuel M. , YORK Michele K. , MONTAGUE P. Read. The neural substrates of reward processing in humans : the modern role of FMRI -- The Neuroscientist, 10(3):260-268, 2004
McCOY N. L. , PITINO L. Pheromonal influences on sociosexual behavior in young women -- Physiology & Behavior, 75(3):367-375, 2002
MCMILLEN C. , ZURAVIN S. , RIDEOUT G. Perceived benefit from child sexual abuse -- J. Consult Clin. Psychol., 63(6):1037-1043, 1995
MEISEL Robert L. , SACHS Benjamin D. : The physiology of male sexual behavior ? in KNOBIL Ernest , NEILL Jimmy D. : The physiology of reproduction ? Raven Press, 2nd edition, 1994
MEIZNER I. Sonographic observation of in utero fetal "masturbation" -- Journal of Ultrasound in Medicine, 6(2):111, 1987
MENDES-LEITE Rommel : Bisexualité : le dernier tabou ? Calmann Levy 1996
MEREDITH M. Chronic recording of vomeronasal pump activation in awake behaving hamsters -- Physiology & Behavior, 56(2):345-354, 1994
MEREDITH Michael : Human vomeronasal function : a critical review of best and worst cases -- Chemical senses, 26:433-445, 2001
MEREDITH Michael : Sensory processing in the main and accessory olfactory systems : comparisons and contrasts ? Journal of steroid biochemistry and molecular biology, 39/4B:601-614, 1991
MESTON Cindy M. , FROHLICH Penny F. The neurobiology of sexual function -- Archives of General Psychiatry, 57(11):1012-1030, 2000
MICHAEL R. P. , CLANCY A. N. , ZUMPE D. Effects of mating on c-fos expression in the brains of male macaques -- Physiology & Behavior, 66(4):591-597, 1999
MICHAEL RP, KEVERNE EB. Pheromones in the communication of sexual status in primates. Nature 1968; 218:746–9
MIGNAULT Christine : Les initiatives sexuelles des femelles singes ? La recherche, 293:70-73, 1996
MILLER B. L. , CUMMINGS J. L. , MCINTYRE H. , EBERS G. , GRODE M. Hypersexuality or altered sexual preference following brain injury -- J. Neurol. Neurosurg. Psychiatry, 49(8):867-873, 1986
MINTZ J. , O'HARE K. , O'BRIEN C. P. , GOLDSCHMIDT J. Sexual problems of heroin addicts -- Archives of General Psychiatry, 31(5):700-703, 1974
MIRIN S. M. , MEYER R. E. , MENDELSON J. H. , ELLINGBOE J. Opiate use and sexual function -- The American journal of psychiatry, 137(8):909-915, 1980
MISSAKIAN E. A. Reproductive behavior of socially deprived male rhesus monkeys (Macaca mulatta) -- J. Comp Physiol Psychol., 69(3):403-407, 1969
MOBBS Dean , GREICIUS Michael D. , ABDEL-AZIM Eiman , MENON Vinod , REISS Allan L. Humor modulates the mesolimbic reward centers -- Neuron, 40(5):1041-1048, 2003
MONCHO-BOGANI J. , LANUZA E. , HERNANDEZ A. , NOVEJARQUE A. , MARTINEZ-GARCIA F. Attractive properties of sexual pheromones in mice: innate or learned? -- Physiology & Behavior, 77(1):167-176, 2002
MONCHO-BOGANI J. , LANUZA E. , LORENTE M. J. , MARTINEZ-GARCIA F. Attraction to male pheromones and sexual behaviour show different regulatory mechanisms in female mice -- Physiology & Behavior, 81(3):427-434, 2004
MONCHO-BOGANI J. , MARTINEZ-GARCIA F. , NOVEJARQUE A. , LANUZA E. Attraction to sexual pheromones and associated odorants in female mice involves activation of the reward system and basolateral amygdala -- Eur. J. Neurosci., 21(8):2186-2198, 2005
MONEY J. Use of androgen-depleting hormone in the treatment of male sex offenders -- Journal of Sex Research, 6:165-172, 1970
MONEY John : Paraphilias : phenomenology and classification ? American journal of psychotherapy, 38/2:164-179, 1984
MONEY John : The conceptual neutering of gender and the criminalization of sex ? Archives of sexual behavior, 14/3:279-290, 1985
MONTAGU A. The skin, touch, and human development. Clinics in Dermatology, 2/4:17-26, 1984
MONTAGU Ashley. La peau et le toucher, un premier langage. Seuil 1979
MONTAGUE P. R. , HYMAN S. E. , COHEN J. D. Computational roles for dopamine in behavioural control -- Nature, 431(7010):760-767, 2004
MOORE C. L. Maternal contributions to the development of masculine sexual behavior in laboratory rats -- Dev. Psychobiol., 17(4):347-356, 1984
MOORE C. L. The role of maternal stimulation in the development of sexual behavior and its neural basis -- Annals of the New York Academy of Sciences, 662:160-177, 1992
MORRIS Desmond. Intimate behavior · Londres, 1971 et Paris, Grasset, 1972
MORRIS Desmond. Patterns of reproductive behaviour · Jonathan Cape, London, 1970
MORRIS J. A. , JORDAN C. L. , BREEDLOVE S. M. Sexual differentiation of the vertebrate nervous system -- Nature Neuroscience, 7(10):1034-1039, 2004
MORRIS J. S. , FRITH C. D. , PERRETT D. I. , ROWLAND D. , YOUNG A. W. , CALDER A. J. , DOLAN R. J. A differential neural response in the human amygdala to fearful and happy facial expressions -- Nature, 383(6603):812-815, 1996
MOSES Jason , LOUCKS Julie A. , WATSON Heather L. , MATUSZEWICH Leslie , HULL Elaine M. : Dopaminergic drugs in the medial preoptic area and nucleus accumbens : effects on motor activity, sexual motivation, and sexual performance ? Pharmacology biochemistry and behavior, 51/4:681-686, 1995
MOSSUZ-LAVAU Janine. La vie sexuelle en France -- La Martinière, 2002
MOURAS Harold , STOLERU Serge , BITTOUN Jacques , GLUTRON D. , PELEGRINI-ISSAC M. , PARADIS A. L. , BURNOD Y. Brain processing of visual sexual stimuli in healthy men : a functional magnetic resonance imaging study -- Neuroimage, 20(2):855-869, 2003
NADAL Jean-Pierre. Réseaux de neurones : de la physique à la psychologie. Armand Colin, 1993
NAKAKUKI Masafumi. Normal and developmental aspects of masochism: transcultural and clinical implications -- Psychiatry, 57(3):244-257, 1994
NAKAMURA K. , KAWASHIMA R. , ITO K. , SUGIURA M. , KATO T. , NAKAMURA A. , HATANO K. , NAGUMO S. , KUBOTA K. , FUKUDA H. , KOJIMA S. Activation of the right inferior frontal cortex during assessment of facial emotion -- Journal of Neurophysiology, 82(3):1610-1614, 1999
NANDA Serena : The Hijras of India : cultural and individual dimensions of an institutionalized third gender role ? in BLACKWOOD Evelyne (Ed) : The many faces of homosexuality : anthropological approaches to homosexual behavior ? Harrington Park Press, New York, 1986
NESTLER E. J. , AGHAJANIAN G. K. Molecular and cellular basis of addiction -- Science, 278(5335):58-63, 1997
NICOLI R.M. , NICOLI J.M. : Biochimie de l'éros ? Contracept. Fertil. Sex., 23/2:137-144, 1995
NIEUWENHUYS R. , VOOGD J. , VAN HUIJZEN, Chr. : The human nervous system : a synopsis and atlas -- Springer-verlag, 3rd edition, 1988
NORMAN Reid L. , SPIES Harold G. : Cyclic oavarian function in a male macaque : additionnal evidence for a lack of sexual differenciation in the physiological mechanisms that regulate the cyclic release of gonadotrophins in primates ? Endocrinology, 118/6:2608-2610, 1986
OATLEY Keith , JENKINS Jennifer M. : Human emotion : function and dysfunction · Annu. rev. psychol., 43:55-85, 1992
O'CONNELL H. E. , DELANCEY J. O. Clitoral anatomy in nulliparous, healthy, premenopausal volunteers using unenhanced magnetic resonance imaging -- The Journal of Urology, 173(6):2060-2063, 2005
O'CONNELL H. E. , HUTSON J. M. , ANDERSON C. R. , PLENTER R. J. Anatomical relationship between urethra and clitoris -- The Journal of Urology, 159(6):1892-1897, 1998
ODENDAAL J. S. Animal-assisted therapy - magic or medicine? -- Journal of Psychosomatic Research, 49(4):275-280, 2000
O'DOHERTY J. , ROLLS Edmund T. , FRANCIS S. , BOWTELL R. , MCGLONE F. Representation of pleasant and aversive taste in the human brain -- Journal of Neurophysiology, 85(3):1315-1321, 2001
O'DWYER A. M. , MARKS I. Obsessive-compulsive disorder and delusions revisited -- Br. J. Psychiatry, 176:281-284, 2000
OKAMI Paul , OLMSTEAD Richard , ABRAMSON Paul R. Sexual experiences in early childhood : 18-year longitudinal data from the UCLA family lifestyles project -- The Journal of Sex Research, 34(4):339-347, 1997
OKAMI Paul , OLMSTEAD Richard , ABRAMSON Paul R. , PENDLETON Laura. Early childhood exposure to parental nudity and scenes of parental sexuality ("primal scenes"): an 18-year longitudinal study of outcome -- Archives of Sexual Behavior, 27(4):361-384, 1998
OLAUSSON H. , LAMARRE Y. , BACKLUND H. , MORIN C. , WALLIN B. G. , STARCK G. , EKHOLM S. , STRIGO I. , WORSLEY K. , VALLBO A. B. , BUSHNELL M. C. Unmyelinated tactile afferents signal touch and project to insular cortex -- Nature Neuroscience, 5(9):900-904, 2002
OLDENBURGER W. P. , EVERITT B. J. , DE JONGE F. H. Conditioned place preference induced by sexual interaction in female rats -- Hormones and Behavior, 26(2):214-228, 1992
OLSEN Kathie L. : Genetic influences on sexual behavior differentiation ? in GERAL Arnold A. , MOLTZ Howard , WARD Ingeborg L. (Ed) : Sexual differentiation, vol 11, Handbook of behavioral neurobiology ? Plenum Press, NY, 1992
OOMURA Y. , YOSHIMATSU H. , AOU S. : Medial preoptic and hypothalamic neuronal activity during sexual behavior of the male monkey ? Brain research, 266:340-343, 1983
PANDI-PERUMAL S. R. , SRINIVASAN V. , MAESTRONI G. J. , CARDINALI D. P. , POEGGELER B. , HARDELAND R. Melatonin: Nature's most versatile biological signal? -- FEBS J., 273(13):2813-2838, 2006
PANKSEPP Jaak , BURGDORF Jeff. "Laughing" rats and the evolutionary antecedents of human joy? -- Physiology & Behavior, 79(3):533-547, 2003
PANKSEPP Jaak , HERMAN B. H. , VILBERG T. , BISHOP P. , DEESKINAZI F. G. Endogenous opioids and social behavior -- Neuroscience and Biobehavioral Reviews, 4(4):473-487, 1980
PANKSEPP Jaak , KNUTSON B. , BURGDORF Jeff. The role of brain emotional systems in addictions: a neuro-evolutionary perspective and new 'self-report' animal model -- Addiction, 97(4):459-469, 2002
PANKSEPP Jaak : A critical role for "affective neuroscience" in resolving what is basis about basic emotions · Psychological review, 99/3:554-560, 1992
PANKSEPP Jaak : Affective neuroscience : the foundations of human and animal emotions · Oxford University Press 1998
PAPINI Mauricio R. Comparative psychology of surprising nonreward -- Brain, Behavior and Evolution, 62(2):83-95, 2003
PARADISO S. , JOHNSON D. L. , ANDREASEN N. C. , O'LEARY D. S. , WATKINS G. L. , PONTO L. L. , HICHWA R. D. Cerebral blood flow changes associated with attribution of emotional valence to pleasant, unpleasant, and neutral visual stimuli in a PET study of normal subjects -- The American journal of psychiatry, 156(10):1618-1629, 1999
PAREDES R. G. Medial preoptic area/anterior hypothalamus and sexual motivation -- Scandinavian Journal of Psychology, 44(3):203-212, 2003
PAREDES R. G. , AGMO Anders. Has dopamine a physiological role in the control of sexual behavior ? A critical review of the evidence -- Progress in Neurobiology, 73(3):179-226, 2004
PAREDES R. G. , ALONSO A. Sexual behavior regulated (paced) by the female induces conditioned place preference -- Behavioral Neuroscience, 111(1):123-128, 1997
PARKER A. J. , NEWSOME W. T. Sense and the single neuron: probing the physiology of perception -- Annu. Rev. Neurosci., 21:227-277, 1998
PARTRIDGE Burgo. A history of orgies -- Prion, 2002 (première publication 1958)
PAUGAM-MOISY Hélène, ROYET Jean-Pierre, ZIGHED Djamel A. Le neuromimétisme : épistémologie, neurobiologie, informatique. Hermes, 1995
PECINA S. , BERRIDGE K. C. Central enhancement of taste pleasure by intraventricular morphine -- Neurobiology (Bp), 3(3-4):269-280, 1995
PEDREIRA D. A. , YAMASAKI A. , CZERESNIA C. E. Fetal phallus 'erection' interfering with the sonographic determination of fetal gender in the first trimester -- Ultrasound in Obstetrics and Gynecology, 18(4):402-404, 2001
PELLETIER Mario , BOUTHILLIER Alain , LEVESQUE Johanne , CARRIER Serge , BREAULT Claude , PAQUETTE Vincent , MENSOUR Boualem , LEROUX Jean-Maxime , BEAUDOIN Gilles , BOURGOUIN Pierre , BEAUREGARD Mario. Separate neural circuits for primary emotions? Brain activity during self-induced sadness and happiness in professional actors -- Neuroreport, 14(8):1111-1116, 2003
PELUSO Emanuel , PUTNAM Nicholas : Case study : sexual abuse of boys by females ? J. Am. Acad. Child Adolesc. Psychiatry, 35/1:51-54, 1996
PEREZ-MARRERO R. A sexuality survey of young adults with spina-bifida -- The Journal of Urology, 131:104A, 1984
PETERSEN S. E. , VAN Mier H. , FIEZ J. A. , RAICHLE M. E. The effects of practice on the functional anatomy of task performance -- Proceedings of the National Academy of Sciences of the United States of America, 95(3):853-860, 1998
PFAFF Donald W. , SCHWARTZ-GIBLIN Susan , MACCARTHY Margareth M. , KOW Lee-Ming : Cellular and molecular mechanisms of female reproductive behaviors ? in KNOBIL Ernest , NEILL Jimmy D. : The physiology of reproduction ? Raven Press, 2nd edition, 1994
PFAUS J. G. , KIPPIN T. E. , CENTENO S. Conditioning and sexual behavior: a review -- Hormones and Behavior, 40(2):291-321, 2001
PFAUS J. G. , KIPPIN T. E. , CORIA-AVILA G. What can animal models tell us about human sexual response? -- Annu. Rev. Sex Res., 14:1-63, 2003
PFAUS James G. Neurobiology of sexual behavior -- Current Opinion in Neurobiology, 9(6):751-758, 1999
PITHERS W. D. , GRAY A. , BUSCONI A. , HOUCHENS P. Children with sexual problems : identification of five distinct child types and related treatment considerations -- Child maltreatment, 3(4):384-406, 1998
PLOOG Detlev W. The place of the Triune Brain in psychiatry -- Physiology & Behavior, 79(3):487-493, 2003
PLUMMER Ken : Understanding childhood sexualities ? Journal of homosexuality, 20/1-2:231-249, 1990
POLLACK Michael : L'homosexualité masculine, ou : le bonheur dans le ghetto ? ? Communications, 35:37-55, 1982
POPPER Karl R. : Le réalisme et la science · Hermann 1990
PORGES Stephen W. Love : an emergent property of the mammalian autonomic nervous system -- Psychoneuroendocrinology, 23(8):837-861, 1998
PORTER Richard H. Olfaction and human kin recognition -- Genetica, 104(3):259-263, 1999
PORTER Richard H. , MOORE John D. : Human kin recognition by olfactory cues ? Physiology and behavior, 27/3:493-495, 1981
POUDAT François-Xavier , JARROUSSE Noëlla : Traitement comportemental et cognitif des difficultés sexuelles ? Masson, 2e édition, 1992
PRESCOTT James W. : Body pleasure and the origins of violence ? Futurist, april, p64-74, 1975
PRETI G. , CUTLER W. B. , CHRISTENSEN C. M. , LAWLEY H. , HUGGINS G. R. , GARCIA C.-R. Human axillary extracts: Analysis of compounds from samples which influence menstrual timing -- Journal of Chemical Ecology, 13(4):717-732, 1987
PURVES Dale , AUGUSTINE George J. , FITZPATRICK David , KATZ Lawrence C. , LAMANTIA Antony-Samuel , MCNAMARA James O. , WILLIAMS Mark S. : Le sexe, la sexualité et le cerveau · in Neuroscience · De Boeck Université, 2nd édition, 2003, p645-664
RAMPIN O. , GIULIANO F. [Physiology and pharmacology of ejaculation] -- J. Soc. Biol., 198(3):231-236, 2004
RAMPIN Olivier , GIULIANO François , BENOIT Gérard , JARDIN Alain : Commande nerveuse centrale de l'érection ? Progrès en urologie, 7:17-23, 1997
RAMPIN Olivier. Sexe et cerveau : quel est l'apport de la recherche fondamentale ? -- Sexologies, XIII(49):29-33, 2004
RAUCH S. L. , SHIN L. M. , DOUGHERTY D. D. , ALPERT N. M. , ORR S. P. , LASKO M. , MACKLIN M. L. , FISCHMAN A. J. , PITMAN R. K. Neural activation during sexual and competitive arousal in healthy men -- Psychiatry Research, 91(1):1-10, 1999
RAUCH S. L. , SHIN L. M. , WRIGHT C. I. Neuroimaging studies of amygdala function in anxiety disorders -- Annals of the New York Academy of Sciences, 985:389-410, 2003
RAUSCHER F. H. , SHAW G. L. , KY K. N. Listening to Mozart enhances spatial-temporal reasoning: towards a neurophysiological basis -- Neuroscience Letters, 185(1):44-47, 1995
RAUSCHER F. H. , SHAW G. L. , KY K. N. Music and spatial task performance -- Nature, 365(6447):611, 1993
RAWN David J. : Traité de biochimie · Éditions Universitaires 1990
REDON Richard & al. Global variation in copy number in the human genome -- Nature, 444(7118):444-454, 2006
REDOUTE Jérôme , STOLERU Serge , GREGOIRE Marie-Claude , COSTES N. , CINOTTI L. , LAVENNE F. , LE Bars D. , FOREST M. G. , PUJOL J. F. Brain processing of visual sexual stimuli in human males -- Human Brain Mapping, 11(3):162-177, 2000
REICH Wilhelm : La révolution sexuelle ? Plon, 10/18, 1968
REIMAN Eric M. The application of positron emission tomography to the study of normal and pathologic emotions -- J. Clin. Psychiatry, 58 Suppl 16:4-12, 1997
REIMAN Eric M. , LANE R. D. , AHERN G. L. , SCHWARTZ G. E. , DAVIDSON R. J. , FRISTON K. J. , YUN L. S. , CHEN K. Neuroanatomical correlates of externally and internally generated human emotion -- The American journal of psychiatry, 154(7):918-925, 1997
REINISCH June Machover , SANDERS Stephanie A. : Prenatal hormonal contributions to sex differences in human cognitive and personality development ? in GERAL Arnold A. , MOLTZ Howard , WARD Ingeborg L. (Ed) : Sexual differentiation, vol 11, Handbook of behavioral neurobiology ? Plenum Press, NY, 1992
RÉMÈS Érik : Sexe guide · Éditions blanche 2004
REPPUCCI N. D. , HAUGAARD J. J. Prevention of child sexual abuse. Myth or reality -- The American Psychologist, 44(10):1266-1275, 1989
REYNAUD Michel. L'amour est une drogue douce ... en général -- Robert Laffont, 2005
RICHARD-FOY Christian : Communication particulière. 2004
RIEGER G. , CHIVERS M. L. , BAILEY J. M. Sexual arousal patterns of bisexual men -- Psychol. Sci., 16(8):579-584, 2005
RILLING James K. , SANFEY Allan G. , ARONSON Jessica A. , NYSTROM Leigh E. , COHEN Jonathan D. The neural correlates of theory of mind within interpersonal interactions -- Neuroimage, 22(4):1694-1703, 2004
RISCH Neil : Male sexual orientation and genetic evidence ? Science, 262:2063-2065, 1993
ROBINSON Terry E. , BERRIDGE Kent C. Addiction -- Annual Review of Psychology, 54:25-53, 2003
ROCHE Bruno , SKALA K. , ROBERT-YAP J. Sexualité et proctologie -- Sexologies, XIII(49):21-25, 2004
RODRIGUEZ-RUST Paula C. Bisexuality : the state of the union -- Annual Review of Sex Research, 13:180-240, 2002
ROHEN Johannes W. , YOKOCHI Chihiro : Anatomie humaine : atlas photographique de l'anatomie systématique et topographique · Éditions Vigot, Paris, 1991
ROLLS Edmund T. , KRINGELBACH Morten L. , DE ARAUJO Ivan E. T. Different representations of pleasant and unpleasant odours in the human brain -- Eur. J. Neurosci., 18(3):695-703, 2003
ROLLS Edmund T. , O'DOHERTY J. , KRINGELBACH Morten L. , FRANCIS S. , BOWTELL R. , MCGLONE F. Representations of pleasant and painful touch in the human orbitofrontal and cingulate cortices -- Cerebral Cortex, 13(3):308-317, 2003
ROSE James D. : Forebrain influences on brainstem and spinal mechanisms of copulatory behavior : a current perspective on Frank Beach's contribution ? Neuroscience and biobehavioral reviews, 14:207-215, 1990
ROSE Robert D. , COLLINS William F., III. Crossing dendrites may be a substrate for synchronized activation of penile motoneurons -- Brain Research, 337(2):373-377, 1985
ROSEN R. C. , LANE R. M. , MENZA M. Effects of SSRIs on sexual function : a critical review -- Journal of Clinical Psychopharmacology, 19(1):67-85, 1999
ROSENZWEIG Mark R. , BREEDLOVE S. Marc , LEIMAN Arnold L. : Biological psychology : an introduction to behavioral, cognitive, and clinical neuroscience -- Sinauer Associates, 3nd edition, 2002
ROSENZWEIG Mark R. , LEIMAN Arnold L. : Psychophysiologie -- Interéditions 1991
ROSSIAUD Jacques : La prostitution dans les villes française au XVe siècle ? Communications, 35:68-84, 1982
ROUBERGUE Anne. Pour éviter le nanisme psychosocial. Cerveau et Psycho, 3:56-59, 2003
ROUBLEV Anastasia. L'identité sexuelle serait déterminée in utéro -- Sexologies, XIII(49):56, 2004
RUSSELL R.J.H. ; WELLS P.A. , RUSHTON J.P. : Evidence for genetic similarity detection in human marriage ? Ethology and sociobiology, 6:183-187, 1985
SACHS Benjamin D. : Penile erection in response to remote cues from females : albino rats severely impaired relative to pigmented strains ? Physiology and behavior, 60/3:803-808, 1996
SACHS Benjamin D. : Placing erection in context : the reflexogenic-psychogenic dichotomy reconsidered ? Neuroscience and biobehavioral reviews, 19:211-224, 1995
SACHSER Norbert , DURSCHLAG Matthias , HIRZEL Daniela. Social relationships and the management of stress -- Psychoneuroendocrinology, 23(8):891-904, 1998
SANFEY Allan G. , RILLING James K. , ARONSON Jessica A. , NYSTROM Leigh E. , COHEN Jonathan D. The neural basis of economic decision-making in the Ultimatum Game -- Science, 300(5626):1755-1758, 2003
SAPER Clifford B. , CHOU Thomas C. , ELMQUIST Joel K. The need to feed : homeostatic and hedonic control of eating -- Neuron, 36(2):199-211, 2002
SARGENT Peter B. : Signalisation électrique · in HALL Zach W. : Introduction à la neurobiologie moléculaire · Médecine-Sciences / Flammarion, 1994
SAVIC I. , BERGLUND H. , LINDSTROM P. Brain response to putative pheromones in homosexual men -- Proceedings of the National Academy of Sciences of the United States of America, 102(20):7356-7361, 2005
SAVIC I. , BERGLUND H. , LINDSTROM P. Brain response to putative pheromones in homosexual men -- Proceedings of the National Academy of Sciences of the United States of America, 102(20):7356-7361, 2005
SAVIC Ivanka , BERGLUND Hans , GULYAS Balazs , ROLAND Per. Smelling of odorous sex hormone-like compounds causes sex-differentiated hypothalamic activations in humans -- Neuron, 31(4):661-668, 2001
SAVIC Ivanka , BERGLUND Hans. Passive perception of odors and semantic circuits -- Human Brain Mapping, 21(4):271-278, 2004
SAVIC Ivanka , GULYAS Balazs , BERGLUND Hans. Odorant differentiated pattern of cerebral activation : comparison of acetone and vanillin -- Human Brain Mapping, 17(1):17-27, 2002
SAVIC Ivanka. Brain imaging studies of the functional organization of human olfaction -- The Neuroscientist, 8(3):204-211, 2002
SCHAAL B. , COUREAUD G. , LANGLOIS D. , GINIES C. , SEMON E. , PERRIER G. Chemical and behavioural characterization of the rabbit mammary pheromone -- Nature, 424(6944):68-72, 2003
SCHAAL B. : Olfaction et processus sociaux chez l'homme : Bref bilan ? Revue internationale de psychopathologie (Paris), 22:387-421, 1996
SCHLAEPFER T. E. , STRAIN E. C. , GREENBERG B. D. , PRESTON K. L. , LANCASTER E. , BIGELOW G. E. , BARTA P. E. , PEARLSON G. D. Site of opioid action in the human brain: mu and kappa agonists' subjective and cerebral blood flow effects -- The American journal of psychiatry, 155(4):470-473, 1998
SCHLESINGER Louis B. Adolescent sexual matricide following repetitive mother-son incest -- Journal of Forensic Science, 44(4):746-749, 1999
SCHMIDT Gunter. The dilemma of the male pedophile -- Archives of Sexual Behavior, 31(6):473-477, 2002
SCHREIDER Eugène : Les unions consanguines : mythes, réalités, tabous ? La recherche, 90:544-551, 1978
SCHUILING G. A. The benefit and the doubt: why monogamy? -- J. Psychosom. Obstet. Gynaecol., 24(1):55-61, 2003
SCHULTZ Willibrord W. , VAN ANDEL Pek , SABELIS Ida , MOOYAART Eduard. Magnetic resonance imaging of male and female genitals during coitus and female sexual arousal -- BMJ, 319(7225):1596-1600, 1999
SCHULTZ Wolfram. Neural coding of basic reward terms of animal learning theory, game theory, microeconomics and behavioural ecology -- Current Opinion in Neurobiology, 14(2):139-147, 2004
SCHWARTZ C. E. , WRIGHT C. I. , SHIN L. M. , KAGAN J. , RAUCH S. L. Inhibited and uninhibited infants "grown up": adult amygdalar response to novelty -- Science, 300(5627):1952-1953, 2003
SEGRAVES Robert T. Antidepressant-induced orgasm disorder -- Journal of Sex and Marital Therapy, 21(3):192-201, 1995
SEGRAVES Robert T. Antidepressant-induced sexual dysfunction -- J. Clin. Psychiatry, 59 Suppl 4:48-54, 1998
SELDEN Samuel T. Tickle -- Journal of the American Academy of Dermatology, 50(1):93-97, 2004
SELL L. A. , MORRIS J. , BEARN J. , FRACKOWIAK R. S. , FRISTON K. J. , DOLAN R. J. Activation of reward circuitry in human opiate addicts -- Eur. J. Neurosci., 11(3):1042-1048, 1999
SERGENT Bernard : L'homosexualité initiatique dans l'europe ancienne ? Payot 1986
SERON Xavier , JEANNEROD Marc ; (Ed) : Neuropsychologie humaine · Mardaga, Liège, 1994
SEWARDS Terence V. Dual separate pathways for sensory and hedonic aspects of taste -- Brain Research Bulletin, 62(4):271-283, 2004
SEWARDS Terence V. , SEWARDS Mark A. Representations of motivational drives in mesial cortex, medial thalamus, hypothalamus and midbrain -- Brain Research Bulletin, 61(1):25-49, 2003
SHEEHAN T. P. , CHAMBERS R. A. , RUSSELL D. S. Regulation of affect by the lateral septum : implications for neuropsychiatry -- Brain Research : Brain Research Reviews, 46(1):71-117, 2004
SHEPHER Joseph : Mate selection among second generation kibbutz adolescents and adults : incest avoidance and negative imprinting ? Archives of sexual behavior, 1/4:293-307, 1971
SHIMA K. , TANJI J. Role for cingulate motor area cells in voluntary movement selection based on reward -- Science, 282(5392):1335-1338, 1998
SHIMURA T. , YAMAMOTO T. , SHIMOKOCHI M. The medial preoptic area is involved in both sexual arousal and performance in male rats: re-evaluation of neuron activity in freely moving animals -- Brain Research, 640(1-2):215-222, 1994
SHIPLEY M.T. , MURPHY A.Z. , RIZVI T.A. , ENNIS M. , BEHBEHANI M.M. : Olfaction and brain stem circuits of reproductive behavior in the rat ? in HOLSTEGE G. , BANDLER R. , SAPER C.B. : The emotional motor system ? Elsevier 1996
SHIROZU H. , KOYANAGI T. , TAKASHIMA T. , HORIMOTO N. , AKAZAWA K. , NAKANO H. Penile tumescence in the human fetus at term--a preliminary report -- Early Hum. Dev., 41(3):159-166, 1995
SIGNORET Jean-Pierre , BALTHAZART Jacques : Le comportement sexuel ? in THIBAULT Charles ; LEVASSEUR Marie-claire : La reproduction chez les mammifères et l'homme ? INRA Ellipse 1991
SIGNORET Jean-Pierre : Olfaction et sexualité des mammifères ? Revue internationale de psychopathologie, 22:423-443, 1996
SIGNORET Jean-Pierre : Sexuel (Comportement) ? Encyclopædia Universalis, version CD-ROM 5.0 (1999)
SIGUSCH V. [Social transformation of sexuality in the past decades. An overview] -- Fortschr. Neurol. Psychiatr., 68(3):97-106, 2000
SIMON Victor. Abus sexuels sur mineurs -- Armand Colin, 2004
SIPSKI Marca L. Sexual functioning in the spinal cord injured -- Int. J. Impot. Res., 10 Suppl 2:S128-S130, 1998
SIPSKI Marca L. Sexual response in women with spinal cord injury: neurologic pathways and recommendations for the use of electrical stimulation -- J. Spinal Cord. Med., 24(3):155-158, 2001
SIPSKI Marca L. , ALEXANDER Craig J. , ROSEN Raymond. Sexual arousal and orgasm in women : effects of spinal cord injury -- Annals of Neurology, 49(1):35-44, 2001
SISK C. L. , FOSTER D. L. The neural basis of puberty and adolescence -- Nature Neuroscience, 7(10):1040-1047, 2004
SOBEL Noam , BROWN Windy M. The scented brain : pheromonal responses in humans -- Neuron, 31(4):512-514, 2001
SOULIER Bernadette. Un amour comme tant d'autres ? Handicaps moteurs et sexualité -- Association des paralysés de France, 2001
SPECTOR Ilana P. , CAREY Michael P. Incidence and prevalence of the sexual dysfunctions: a critical review of the empirical literature -- Archives of Sexual Behavior, 19(4):389-408, 1990
SPECTOR Ilana P. , CAREY Michael P. , STEINBERG Lynne. The sexual desire inventory: development, factor structure, and evidence of reliability -- Journal of Sex and Marital Therapy, 22(3):175-190, 1996
SPEVAK A. M. , QUADAGNO D. M. , KNOEPPEL D. The effects of isolation on sexual and social behavior in the rat -- Behav. Biol., 8(1):63-73, 1973
SPIES H. G. , PAU K. Y. , YANG S. P. Coital and estrogen signals: a contrast in the preovulatory neuroendocrine networks of rabbits and rhesus monkeys -- Biol. Reprod., 56(2):310-319, 1997
SPITZ R. Hospitalism. An inquiry into the genesis of psychiatric conditions in early childhood. Psychoanalytic Study of the Child, 1:53-174, 1945
STELLAR James R. : Investigating the neural circuitry of brain stimulation reward · Progress in psychobiology and physiological psychology, 14:235-294, 1990
STENSAAS Larry J. , LAUKER Robert M. , MONTI-BLOCH L. , GROSSER Bernard I. , BERLINER David L. : Ultrastructure of the human VNO ? Journal of steroid biochemistry and molecular biology, 39:553-560, 1991
STERN J. M. Offspring-induced nurturance: animal-human parallels -- Dev. Psychobiol., 31(1):19-37, 1997
STERN J. M. , DIX L. , BELLOMO C. , THRAMANN C. Ventral trunk somatosensory determinants of nursing behavior in Norway rats: 2. Role of nipple and surrounding sensations -- Psychobiology, 20(1):71-80, 1992
STERN J. M. , JOHNSON S. K. Ventral somatosensory determinants of nursing behavior in Norway rats. I. Effects of variations in the quality and quantity of pup stimuli -- Physiology & Behavior, 47(5):993-1011, 1990
STERN Judith M. : Multisensory regulation of maternal behavior and masculine sexual behavior : a revised view ? Neuroscience and biobehavioral reviews, 14:183-200, 1990
STOLERU Serge , GREGOIRE Marie-Claude , GERARD Daniel , DECETY Jean , LAFARGE E. , CINOTTI L. , LAVENNE F. , LE Bars D. , VERNET-MAURY E. , RADA H. , COLLET C. , MAZOYER Bernard , FOREST M. G. , MAGNIN F. , SPIRA A. , COMAR D. Neuroanatomical correlates of visually evoked sexual arousal in human males -- Archives of Sexual Behavior, 28(1):1-21, 1999
STOLERU Serge , REDOUTE Jérôme , COSTES N. , LAVENNE F. , BARS D. L. , DECHAUD H. , FOREST M. G. , PUGEAT M. , CINOTTI L. , PUJOL J. F. Brain processing of visual sexual stimuli in men with hypoactive sexual desire disorder -- Psychiatry Research, 124(2):67-86, 2003
STOWERS L. , HOLY T. E. , MEISTER M. , DULAC C. , KOENTGES G. Loss of sex discrimination and male-male aggression in mice deficient for TRP2 -- Science, 295(5559):1493-1500, 2002
STOWERS L. , MARTON T. F. What is a pheromone? Mammalian pheromones reconsidered -- Neuron, 46(5):699-702, 2005
STROHMAN R. C. The coming Kuhnian revolution in biology -- Nat. Biotechnol., 15(3):194-200, 1997
SUGGS Robert C : Marquesan sexual behavior : an anthropological study of polynesian practices · Harcourt, Brace & World, 1966
SUH D. D. , YANG C. C. , CAO Y. , HEIMAN J. R. , GARLAND P. A. , MARAVILLA K. R. MRI of female genital and pelvic organs during sexual arousal -- J. Psychosom. Obstet. Gynaecol., 25(2):153-162, 2004
SUMMIT Roland , KRYSO JoAnn : Sexual abuse of children : a clinical spectrum ? in CONSTANTINE Larry L. , MARTINSON Floyd M. , (Ed) : Children and sex : new findings, new perspectives ? Little, Brown and Co, Boston, 1981
SWAAB D. F. , CHUN W. C. , KRUIJVER F. P. , HOFMAN M. A. , ISHUNINA T. A. Sexual differentiation of the human hypothalamus -- Adv. Exp. Med. Biol., 511:75-100, 2002
SWAAB D. F. , HOFMAN M. A. Sexual differentiation of the human hypothalamus in relation to gender and sexual orientation -- Trends in Neurosciences, 18(6):264-270, 1995
TASSIN Jean-Pol. (Processus renforcement) -- Proceedings of the National Academy of Sciences of the United States of America, 103(19):7476-7481, 2006
THIBAULT Charles , LEVASSEUR Marie-Claire (Eds). La reproduction chez les mammifères et l'Homme -- INRA Ellipse, 2001
THIBAULT Charles ; LEVASSEUR Marie-Claire (Eds). La reproduction chez les mammifères et l'homme -- INRA Ellipse 1991
THINÈS Georges : Instinct · Encyclopaedia Universalis, version CD-ROM 9.0 (2003)
TIIHONEN Jari , KUIKKA Jyrki , KUPILA Jukka , PARTANEN Kaarina , VAINIO Pauli , AIRAKSINEN Juha , ERONEN Markku , HALLIKAINEN Tero , PAANILA Jarmo , KINNUNEN Ilpo , HUTTUNEN Janne : Increase cerebral blood flow of right prefrontal cortex in man during orgasm ? Neuroscience letters, 170:241-243, 1994
TOBET S.A. , FOX T.O. : Sex differences in neuronal morphology influenced hormonally throughout life ? in GERAL Arnold A. , MOLTZ Howard , WARD Ingeborg L. (Ed) : Sexual differentiation, vol 11, Handbook of behavioral neurobiology ? Plenum Press, NY, 1992
TORII Masafuni , KUBO Katsuharu , SASAKI Takashi. Naloxone and initial estrogen action to induce lordosis in ovariectomized rats: the effect of a cut between the septum and preoptic area -- Neuroscience Letters, 195(3):167-170, 1995
TRUITT W. A. , COOLEN L. M. Identification of a potential ejaculation generator in the spinal cord -- Science, 297(5586):1566-1569, 2002
TURNER C. H. , DAVENPORT R. K. , ROGERS C. M. The effect of early deprivation on the social behavior of adolescent chimpanzees -- The American journal of psychiatry, 125(11):1531-1536, 1969
UVNAS-MOBERG Kerstin. Oxytocin may mediate the benefits of positive social interaction and emotions -- Psychoneuroendocrinology, 23(8):819-835, 1998
VAN der Horst, V , HOLSTEGE G. Sensory and motor components of reproductive behavior: pathways and plasticity -- Behavioural Brain Research, 92(2):157-167, 1998
VAN OOYEN A. , VAN PELT J. : Activity-dependent neurite outgrowth and neural network development · in VAN PELT J. , CORNER M.A. , UYLINGS H.B.M. , LOPES DA SILVA F.H. , (Ed) : The self-organizing brain : from growth cones to functional networks · Elsevier 1994
VAN PELT J. , CORNER M.A. , UYLINGS H.B.M. , LOPES DA SILVA F.H. , (Ed) : The self-organizing brain : from growth cones to functional networks · Elsevier 1994
VANDERSCHUREN L. J. , NIESINK R. J. , VAN REE J. M. The neurobiology of social play behavior in rats -- Neuroscience and Biobehavioral Reviews, 21(3):309-326, 1997
VARELA Francisco , THOMPSON Evan , ROSCH Eleanor. L'inscription corporelle de l'esprit : sciences cognitives et expérience humaine -- Seuil, 1993
VASEY P. L. , DUCKWORTH N. Sexual reward via vulvar, perineal, and anal stimulation: a proximate mechanism for female homosexual mounting in Japanese macaques -- Archives of Sexual Behavior, 35(5):523-532, 2006
VAUCLAIR Jacques : La cognition animale · PUF QSJ 1996
VERNIER Philippe : Le développement et l'évolution du système nerveux · Pour la Science, 302:50-56, 2002
VEYNE Paul : L'homosexualité à Rome ? Communications, 35:26-33, 1982
VIGARELLO Georges : Histoire du viol : XVIe-XXe siècle ? Seuil 1998
VINCENT Lucy. Comment devient-on amoureux ? -- Odile Jacob, 2004
VOLBERT Renate : Sexual knowledge of preschool children · in SANDFORT Theo G.M. , RADEMAKERS Jany , (Ed) : Childhood sexuality : normal sexual behavior and development · Journal of psychology & human sexuality, volume 12, numbers 1/2, 2000
WAGER T. D. , RILLING J. K. , SMITH E. E. , SOKOLIK A. , CASEY K. L. , DAVIDSON R. J. , KOSSLYN S. M. , ROSE R. M. , COHEN J. D. Placebo-induced changes in FMRI in the anticipation and experience of pain -- Science, 303(5661):1162-1167, 2004
WAGNER Christine K. , CLEMENS Lynwood G. : Projections of the paraventricular nucleus of the hypothalamus to the sexually dimorphic lumbosacral region of the spinal cord ? Brain research, 539:254-262, 1991
WALLEN K. , PARSONS W. A. Sexual behavior in same-sexed nonhuman primates: is it relevant to understanding human homosexuality? -- Annu. Rev. Sex Res., 8:195-223, 1997
WALLEN Kim : Desire and ability : hormones and the regulation of female sexual behavior ? Neuroscience and biobehavioral reviews, 14:233-241, 1990
WANG Zuoxin , HULIHAN Terrence J. , INSEL Thomas R. Sexual and social experience is associated with different patterns of behavior and neural activation in male prairie voles -- Brain Research, 767(2):321-332, 1997
WANG Zuoxin , YOUNG Larry J. , LIU Yue , INSEL Thomas R. Species differences in vasopressin receptor binding are evident early in development: comparative anatomic studies in prairie and montane voles -- J. Comp Neurol., 378(4):535-546, 1997
WARD Byron O. : Fetal drug exposure and sexual differentiation of males ? in GERAL Arnold A. , MOLTZ Howard , WARD Ingeborg L. (Ed) : Sexual differentiation, vol 11, Handbook of behavioral neurobiology ? Plenum Press, NY, 1992
WARD Ingeborg L. : Sexual behavior : the product of perinatal hormonal and prepubertal social factors ? in GERAL Arnold A. , MOLTZ Howard , WARD Ingeborg L. (Ed) : Sexual differentiation, vol 11, Handbook of behavioral neurobiology ? Plenum Press, NY, 1992
WEINBERGER Norman M. Dynamic regulation of receptive fields and maps in the adult sensory cortex -- Annu. Rev. Neurosci., 18:129-158, 1995
WEISFELD G. E. , CZILLI T. , PHILLIPS K. A. , GALL J. A. , LICHTMAN C. M. Possible olfaction-based mechanisms in human kin recognition and inbreeding avoidance -- J. Exp. Child Psychol., 85(3):279-295, 2003
WEISNER Thomas S. , WILSON-MITCHELL Jane E. : Nonconventional family life-styles and sex typing in six-year-old ? Child development, 61:1915-1933, 1990
WELLER Leonard , WELLER Aron : Human menstrual synchrony : a critical assessment ? Neuroscience and biobehavioral reviews, 17:427-439, 1993
WESSBERG J. , OLAUSSON H. , FERNSTROM K. W. , VALLBO A. B. Receptive field properties of unmyelinated tactile afferents in the human skin -- Journal of Neurophysiology, 89(3):1567-1575, 2003
WHALEN R. E. Effects of mounting without intromission and intromission without ejaculation on sexual behavior and maze learning -- J. Comp Physiol Psychol., 54:409-415, 1961
WHITE F. J. A behavioral/systems approach to the neuroscience of drug addiction -- The Journal of Neuroscience, 22(9):3303-3305, 2002
WICKENS Jeffery R. , REYNOLDS John N. , HYLAND Brian I. Neural mechanisms of reward-related motor learning -- Current Opinion in Neurobiology, 13(6):685-690, 2003
WILCOX A. J. , BAIRD D. D. , DUNSON D. B. , MCCONNAUGHEY D. R. , KESNER J. S. , WEINBERG C. R. On the frequency of intercourse around ovulation: evidence for biological influences -- Hum. Reprod., 19(7):1539-1543, 2004
WILD Barbara , RODDEN Frank A. , GRODD Wolfgang , RUCH Willibald. Neural correlates of laughter and humour -- Brain, 126(Pt 10):2121-2138, 2003
WILLIS W.D. , COGGESHALL R.E. : Sensory mechanisms of the spinal cord · Plenum Press, NY, 2nd edition, 1991
WILLIS W.D. , WESTLUND K.N. : Neuroanatomy of the pain system and of the pathways that modulate pain · Journal of clinical neurophysiology, 14/1:2-31, 1997
WILSON Gisela F. : Cellular Neuromodulation · Encyclopedia of Life Sciences, Nature Publishing Group, www.els.net, 1999
WILSON H. C. , KIEFHABER S. H. , GRAVEL V. Two studies of menstrual synchrony: negative results -- Psychoneuroendocrinology, 16(4):353-359, 1991
WILSON Mark E. : Factors determining the onset of puberty ? in GERAL Arnold A. , MOLTZ Howard , WARD Ingeborg L. (Ed) : Sexual differentiation, vol 11, Handbook of behavioral neurobiology ? Plenum Press, NY, 1992
WINBERG J. , PORTER Richard H. Olfaction and human neonatal behaviour : clinical implications -- Acta Paediatrica, 87(1):6-10, 1998
WINKELMANN R. K. The cutaneous innervation of human newborn prepuce -- J. Invest Dermatol., 26(1):53-67, 1956
WINKELMANN R. K. The erogenous zones: their nerve supply and its significance -- Mayo Clin. Proc., 34(2):39-47, 1959
WISE R. A. Forebrain substrates of reward and motivation -- J. Comp Neurol., 493(1):115-121, 2005
WOLPE P. R. Ethics and social policy in research on the neuroscience of human sexuality -- Nature Neuroscience, 7(10):1031-1033, 2004
WOODSON James C. Including 'learned sexuality' in the organization of sexual behavior -- Neuroscience and Biobehavioral Reviews, 26(1):69-80, 2002
WUNSCH Serge , BRENOT Philippe. Analyse des rapports entre structure biologique et sexualité -- NeuroPsy News, 4(4):133-136, 2005
WUNSCH Serge , BRENOT Philippe. Does a sexual instinct exist ? -- Sexologies, 13(48):30-36, 2004
WUNSCH Serge , BRENOT Philippe. Neurobiology of pleasure -- Sexologies, 13(50):17-27, 2004
WUNSCH Serge , BRENOT Philippe. Sexualité : instinct ou apprentissage ? -- Médecine Sexuelle, 1:12-21, 2005
WUNSCH Serge , BRENOT Philippe. Un modèle synthétique des dysfonctions et pathologies sexuelles -- NeuroPsy News, 4(5):162-165, 2005
WUNSCH Serge. Déterminants neurobiologiques du comportement sexuel -- Communication, Congrès de sexologie AIHUS, Marseille, 2006
YAMAMOTO D. , ITO H. , FUJITANI K. Genetic dissection of sexual orientation: behavioral, cellular, and molecular approaches in Drosophila melanogaster -- Neurosci. Res., 26(2):95-107, 1996
YANAGIMOTO Masahiro , HONDA Kazumasa , GOTO Yukio , NEGORO Hideo : Afferents originating from the dorsal penile nerve excite oxytocin cells in the hypothalamic paraventricular nucleus of the rat ? Brain research, 733:292-296, 1996
YATES Alayne. Biologic perspective on early erotic development -- Child and Adolescent Psychiatric Clinics of North America, 13(3):479-496, 2004
YATES Alayne. Eroticized children -- in PERRY M. E. (Ed). Handbook of sexology (vol 7) : Childhood and adolescent sexology -- Elsevier Science, (18):325-334, 1990
YEUNG Nick , COHEN Jonathan D. , BOTVINICK Matthew M. The neural basis of error detection : conflict monitoring and the error-related negativity -- Psychological Review, 111(4):931-959, 2004
YOON H. , ENQUIST L. W. , DULAC C. Olfactory inputs to hypothalamic neurons controlling reproduction and fertility -- Cell, 123(4):669-682, 2005
YOUNG L. J. , WANG Z. The neurobiology of pair bonding -- Nature Neuroscience, 7(10):1048-1054, 2004
YOUNG Larry J. , LIM Miranda M. , GINGRICH Brenden , INSEL Thomas R. Cellular mechanisms of social attachment -- Hormones and Behavior, 40(2):133-138, 2001
YOUNG Larry J. , WINSLOW James T. , NILSEN Roger , INSEL Thomas R. Species differences in V1a receptor gene expression in monogamous and nonmonogamous voles: behavioral consequences -- Behavioral Neuroscience, 111(3):599-605, 1997
ZALEL Y. , PINHAS-HAMIEL O. , LIPITZ S. , MASHIACH S. , ACHIRON R. The development of the fetal penis--an in utero sonographic evaluation -- Ultrasound in Obstetrics and Gynecology, 17(2):129-131, 2001
ZAVIACIC M. , ABLIN R. J. The female prostate -- J. Natl. Cancer Inst., 90(9):713-714, 1998
ZHANG J. , WEBB D. M. Evolutionary deterioration of the vomeronasal pheromone transduction pathway in catarrhine primates -- Proceedings of the National Academy of Sciences of the United States of America, 100(14):8337-8341, 2003
ZHOU Jiang-Ning , HOFMAN Michel A. , GOOREN Louis J. , SWAAB Dick F. A sex difference in the human brain and its relation to transsexuality -- Nature, 378(6552):68-70, 1995
ZIMMER-GEMBECK M. J. The development of romantic relationships and adaptations in the system of peer relationships -- Journal of Adolescent Health, 31(6 Suppl):216-225, 2002
ZIMMER-GEMBECK M. J. , SIEBENBRUNER J. , COLLINS W. A. A prospective study of intraindividual and peer influences on adolescents' heterosexual romantic and sexual behavior -- Archives of Sexual Behavior, 33(4):381-394, 2004
ANNEXES
– Proposition de définitions
– Le questionnaire "Enquête sur le comportement sexuel"
– Les résultats complémentaires de l'enquête par questionnaire
– Complément : Problèmes relatifs aux
expériences éthiquement réalisables
Annexe I : Proposition de définitions des concepts clés
Comment se traduisent, en terme de structures et de fonctions biologiques, l'inné et l'instinct ? Que doit-on rechercher afin de tester les hypothèses ?
Cette section propose, basées sur les caractéristiques structurelles et fonctionnelles des organismes biologiques, des définitions de l'inné, de l'instinct et du comportement.
Ces définitions, les plus précises possibles, devraient permettre d'identifier de manière non ambiguë les éléments à examiner qui permettront de vérifier l'existence éventuelle de comportements instinctuels.
a – Inné & acquis
La problématique de l'inné et de l'acquis est une problématique ancienne et polémique, mais c'est également une des questions fondamentales de l'étude des organismes vivants : quelle est la part des caractères innés, qui proviennent de l'interaction entre l'expression du génome et les molécules de la cellule primitive, de celles, acquises, qui proviennent de l'influence des différents environnements (chorionique, utérin, écologique, familial et culturel).
Actuellement, cette approche est jugée non pertinente par de nombreux auteurs, car le développement des organismes est perçu comme une interaction réciproque de différents facteurs qui s'inter-influencent, et dont il serait impossible ou sans intérêt de séparer ou d'identifier les facteurs de causalité. Néanmoins, les dernières connaissances en génétique, en biologie du développement et en neurosciences, grâce à, d'une part, l'identification précise des mécanismes et des processus biologiques en cause, et, d'autre part, grâce à l'identification précise des différents environnements et de leurs influences respectives, permettent d'apporter des éléments de précision. Ces données, même si elles ne permettent pas de tout expliciter, démontrent l'existence de caractéristiques qui sont indubitablement de l'ordre de l'inné, et d'autres qui ne peuvent être qu'acquises.
Par exemple, le système nerveux des mammifères présente de nombreuses caractéristiques innées : l'organisation générale du cerveau est toujours la même (systèmes sensoriels, système moteur, système nerveux autonome, hypothalamus, amygdale, néocortex organisé en six couches de neurones et divisé en lobes…) (NIEUWENHUYS 1988). Ces caractéristiques toujours présentes dépendent de mécanismes moléculaires spécifiques, qui peuvent être identifiés et décrits, et dont le fonctionnement explique la permanence des processus innés observés.
À un premier niveau, dans l'œuf fécondé en développement, on peut mettre en évidence des gènes (dans le noyau ou dans les mitochondries) et des molécules informatives qui vont déterminer de nombreux processus cellulaires (régulation de l'expression du génome, synthèse des protéines, division cellulaire, …). Ensuite on peut mettre en évidence des molécules qui déterminent la spécialisation des cellules (cellules de la peau, des os, du système nerveux, …), l'orientation céphalo-caudale de l'organisme ou la création des membres (GILBERT 1996). On peut même mettre en évidence des mécanismes innés de la variabilité, tel l'épissage alternatif des ARN prémessagers, qui permettent des modifications adaptatives de l'organisme face à son environnement. Il est donc possible d'expliquer les effets de l'influence du milieu sur l'organisme en tant que résultat de processus biologiques identifiables – et dont certains sont innés – plutôt qu'en terme d'interactions globales.
En conclusion, il est généralement possible, en détaillant les processus moléculaires, cellulaires, physiologiques ou neurobiologiques, d'identifier des éléments permanents, innés, résultat d'une "programmation" biologique, qui existent toujours et partout, quel que soit l'environnement, tant que ce dernier n'altère pas l'organisme. L'identification de ces éléments biologiques permanents et l'explication des mécanismes et des processus biologiques qui induisent cette permanence permettent de conclure à l'innéité de ces éléments.
Proposition de définition
Quelle est la raison biologique de la nécessité du concept d' "innéité" ?
Les études en biologie du développement ont montré que l'organisme se développe à partir d'informations codées dans l'environnement de la cellule primitive, et à partir d'informations provenant des différents environnements extérieurs.
La question cruciale est de savoir quelle est la part et la dynamique de ces différentes informations – et tout particulièrement des informations de la cellule primitive – dans l'organisation spatiale et temporelle du développement de l'organisme. L'étude de l'information initiale codée dans l'organisme et de son impact sur le développement est un sujet majeur de la biologie du développement.
Les caractéristiques biologiques d'un élément (structurel ou fonctionnel) innés seraient :
– L'information ayant permis le développement de cet élément est contenue dans l'organisme (information génétique et/ou épigénétique).
– L'élément existe à la fin du développement, quelles que soient les caractéristiques des différents environnements extérieurs à l'organisme (tant que ces derniers n'altèrent pas l'organisme).
Le substantif "inné" fait référence à la naissance, étape particulièrement spectaculaire du développement. Mais la sortie du nouveau-né de l'environnement utérin ne marque ni la fin de son développement ni le début ou la fin des influences des différents environnements. La fin du développement, c'est-à-dire la maturité, plutôt que la naissance, devrait être la période de référence du concept d' "innéité".
En fonction des analyses présentées ci-dessus, il est proposé la définition suivante de l'innéité : est "inné" tout élément (structurel ou fonctionnel) dont l'information développementale préexiste dans l'organisme et dont le développement est indépendant de l'influence des différents environnements (du chorion, de l'utérus, du milieu écologique, familial et culturel). Cet élément est toujours présent à la fin du développement, quelles que soient les caractéristiques de ces différents environnements, sauf dans les cas extrêmes où des caractéristiques particulières ont provoqué un développement pathologique (famine, maladies graves, …), qui dépasse les capacités adaptatives de l'organisme (physiologie du jeûne, système immunitaire, …).
b – Instinct
Qu'est-ce que l' "instinct" ? À quelles réalités biologiques correspond-il ? Quelle définition, basée sur des caractéristiques biologiques, pourrait-on proposer ?
Les deux principaux problèmes liés au concept d'instinct sont, d'une part, la question de sa réalité biologique et, d'autre part, l'élaboration d'une définition précise et opérationnelle du terme.
L'instinct est une des explications qui tentent de répondre à une question fondamentale de la biologie animale : quels sont les facteurs qui déterminent et qui sous-tendent les comportements ? Une première réponse, générale, serait que l'instinct correspondrait au développement ou à une organisation particulière d'une structure biologique spécifique, codée par le génome, et qui contrôlerait les comportements adaptatifs d'une espèce. L'instinct désignerait un processus neurobiologique inné qui est à l'origine d'un comportement qui s'exprime en dehors de tout apprentissage. Ou, exprimé autrement, le comportement instinctuel correspondrait, chez les mammifères, à une activation organisée et précablée du système musculaire squelettique par une structure innée et spécifique du système nerveux.
Le concept d' "instinct" désigne un comportement qui est inné, qui existe en dehors de tout apprentissage et de toute expérience.
L'instinct existe-t-il ?
Les observations éthologiques [16] mettent en évidence l'existence de comportements systématiques et stéréotypés, existants en dehors de tout apprentissage et de toute expérience préalable. Les connaissances en génétique (DARNELL 1993), en biologie du développement (GILBERT 1996) et en neurosciences (VAN PELT & al. 1994 ; VERNIER 2002) montrent qu'il existe un génome, des gènes du développement, des molécules de guidage, des structures neurales précablés, c'est-à-dire tout un ensemble de propriétés et de structures qui rendent possible l'existence chez les mammifères de réactions innées. L'ensemble de ces données rend plausible l'hypothèse éthologique et biologique de l'existence d'un "instinct".
Définition du concept d'instinct
Il existe dans la littérature scientifique de nombreuses définitions de la notion d' "instinct" (THINÈS 2003 ; GALLO & COQUERY 1994 p. 388). Le problème est que, suivant la définition utilisée, la notion d'instinct peut recouvrir des réalités différentes. La prise en compte des données éthologiques combinées avec les données neurobiologiques devrait alors permettre une meilleure précision et une plus grande objectivité de la définition.
Les observations éthologiques montrent que les comportements qui sont considérés comme innés peuvent être soit très stéréotypés (tels les réflexes), soit avoir une expression relativement variable (telles les réactions émotionnelles). En outre, les données neurobiologiques nous enseignent qu'il existe des structures neurales "précablées" (KOESTER & O'LEARY 1994) et donc susceptibles de produire des réactions innées stéréotypées, mais également qu'il existe des mécanismes permettant une certaine plasticité neurale (GAZZANIGA 1995, p. 9-122) qui pourrait être à l'origine de réactions innées variables.
La prise en compte de l’ensemble de ces données amène à proposer deux définitions, l’une au sens strict et l’autre au sens large, de la notion d' "instinct" :
Définition au sens strict
La définition stricto sensu correspond, tout en restant compatible avec les données neurobiologiques, à l'archétype de l'instinct : la "programmation" complète, contrôlée et finalisée d'une action ou d'une réaction comportementale.
Une proposition d'une définition de l'instinct, au sens strict, pourrait ainsi être formulée : action ou réaction comportementale, innée, fixe, immédiatement parfaite sans expérience préalable et sans apprentissage, provoquée par des stimuli internes ou externes spécifiques.
Ces réactions sont déterminées par l'existence de structures neurales spécifiquement organisées (récepteurs spécifiques, et/ou synapses spécifiques, et/ou neuromédiateurs spécifiques, et/ou précablage spécifique...), dont la formation est indépendante des influences du milieu extérieur à l'organisme.
L'exemple type de telles réactions instinctives du système nerveux, est représenté par les réflexes.
Par exemple, c'est l'architecture et les connections spécifiques des neurones sensoriels, des interneurones et des neurones moteurs qui permettent le réflexe du retrait de la main à la douleur. Cette séquence motrice instinctuelle est exécutée parfaitement en dehors de tout apprentissage préalable.
Définition au sens large
La définition lato sensu de l'instinct correspond, tout en restant compatible avec les données neurobiologiques, à la configuration minimaliste et aux limites extrêmes de ce qui est inné. Le critère minimaliste de l'innéité serait la réalisation d'une action ou d'une réaction comportementale en l'absence d'expérience.
Une proposition d'une définition de l'instinct, au sens large, pourrait donc être formulée ainsi : action ou réaction comportementale, innée, réalisée sans expérience préalable, mais dont seule la réaction globale est innée et non les détails variables de sa réalisation.
Ces réactions sont provoquées par l'existence de structures neurales dont la macrostructure est génétiquement déterminée, mais dont la microstructure, plastique, dépend des influences de l'environnement.
L'exemple type de telles réactions comportementales instinctives, au sens large, sont les réactions provoquées par les émotions primaires.
Les réactions instinctives de peur en sont un bon exemple. Elles sont commandées par l'amygdale, qui est une structure clé pour cette réaction émotionnelle (DAVIS 1992 ; LE DOUX 1995). L'amygdale existe chez tous les mammifères et son développement est indépendant des influences externes : tous les mammifères ont ainsi des réactions instinctives de peur. Par contre, la microstructure des neurones de l'amygdale (avec des propriétés électrophysiologiques particulières des membranes neuronales, une organisation spécifique des connexions synaptiques, ...) dépend à la fois des influences internes et externes : ce qui permet que les réactions instinctives de peur soient variables en fonction de l'expérience propre de l'animal.
Transcription biologique de l'instinct
L'existence d'un instinct de la reproduction impliquerait la présence dans l'organisme d'éléments biologiques et neurobiologiques spécifiques : gènes spécifiques et/ou hormones spécifiques et/ou organes spécifiques et/ou structures neurales spécifiques… organisés d'une manière telle que les différentes séquences comportementales de la reproduction soient parfaitement initiées, exécutées et contrôlées.
Dans l'hypothèse d'un instinct de la reproduction au sens strict (hypothèse du « tout programmé »), on devrait pouvoir trouver au minimum chez l'Homme les caractéristiques suivantes :
– Une organisation neurale spécifique du comportement de la reproduction (de manière similaire à celles existant spécifiquement pour la vision, l'audition, la respiration ou la régulation cardio-vasculaire). De plus, cette organisation devrait être relativement similaire à celle de tous les autres mammifères (en effet, les observations phylogénétiques montrent que l'organisation des fonctions fondamentales est semblable d'une espèce à l'autre).
– Des émetteurs et des récepteurs innés produisant et détectant des stimuli spécifiques au comportement de reproduction.
Ces éléments (comme par exemple les phéromones sexuelles et l'organe voméronasal (LIMAN 1996)) devant permettre la reconnaissance des partenaires, l'initiation et la poursuite des comportements reproducteurs.
– Des voies nerveuses spécifiques aux signaux des comportements de la reproduction.
– Une ou plusieurs structures neurales locales, spécifiques aux différentes séquences ou réflexes reproducteurs élémentaires (noyaux spécifiques à l'intromission, aux poussées pelviennes, à la fellation, au cunnilingus, au baiser, etc.). Ces éléments neuraux devant permettre le contrôle local de chacune des différentes activités motrices constitutives des comportements de la reproduction : identification du partenaire, positionnement adéquat du corps, séquences oro- ou génito-génitales, caresses érotiques …
– Enfin, une organisation neurale spécifique, plus générale et plus centrale, permettant un contrôle global du comportement de la reproduction.
Cette organisation spécifique devrait, après traitement des stimuli-signaux spécifiques à la reproduction, induire un phénomène cérébral aboutissant aux activités comportementale de la reproduction. Ce phénomène pourrait être soit de type sensorimoteur (déclenchement et contrôle par des stimuli sensoriels spécifiques d'une succession de séquences réflexes aboutissant aux activités reproductrices), soit de type psychique (perception subjective d'une motivation spécifique produisant une forte tendance comportementale à rechercher, par exemple, le plaisir érotique et/ou le contact physique).
Dans l'hypothèse d'une configuration la plus minimale possible (instinct au sens large) permettant un comportement inné, on devrait pouvoir trouver au minimum chez l'Homme les caractéristiques suivantes :
– Des émetteurs et des récepteurs de signaux (internes, olfactifs, auditifs, visuels...) induisant des comportements permettant la reproduction. Dans cette configuration minimale, il serait quand même nécessaire d'avoir des signaux – même s'ils ne sont pas spécifiques – qui permettent, directement ou indirectement, la reconnaissance des partenaires, l'initiation et la poursuite d'un comportement à visée reproductrice.
– Des structures neurales locales et précablées contrôlant la plupart des réflexes et/ou des séquences motrices élémentaires des activités de la reproduction (positionnement du corps, intromission, poussées pelviennes, fellations, cunnilingus, baiser…).
– Une, plusieurs, ou un réseau, de structures produisant un phénomène neural global conduisant à la reproduction, avec la nécessité d’avoir une forte tendance sensorimotrice (drive), et/ou psychique (motivation), à accomplir des actes aboutissant, directement ou indirectement, aux activités reproductrices (ou au minimum au contact physique hétérosexuel).
La mise en évidence d'éléments biologiques de ce type permettrait de conclure à l'existence d'un instinct de la reproduction, contrôlant toutes ou certaines séquences comportementales nécessaires à la réalisation du coït vaginal. L'analyse quantitative et qualitative de ces éléments instinctuels permettrait d'évaluer la part respective des caractères innés, des comportements instinctuels et des apprentissages dans le comportement de reproduction.
Remarque
Pour donner un exemple concret et connu, la lordose, spécifique du comportement de reproduction et existant chez les mammifères inférieurs, précablée au niveau médullaire et mésencéphalique, peut être considérée comme une preuve de l'existence de séquences motrices "instinctuelles". Elle est considérée dans cette étude comme un archétype, mais simple, de ce qui doit être recherché afin de vérifier l'existence de comportements "instinctuels".
g – Comportement
Préalables
Quelles sont les raisons à l'origine du regroupement de certaines actions motrices en comportements distincts et identifiés ? Quelles sont les raisons à l'origine de la focalisation de l'intérêt scientifique sur tel comportement, à l'exclusion des autres ? Par exemple, pourquoi étudie-t-on le comportement de marche, de course ou de saut, mais pas le comportement "giratoire" ? Pourtant les observations d'enfants (et également d'adultes) montrent que leurs comportements moteurs sont très variés, qu'ils marchent, courent et sautent, mais qu'également ils "gyrent" de toutes les façons que les caractéristiques de l'environnement rendent possible : "toupie", manège, tourniquet, etc. Pourquoi étudie-t-on les comportements associés à la peur ou à la communication mais pas les comportements liés au rire ou à la joie ? Pourtant la joie est, tout comme la peur, considéré comme une émotion primaire, et les comportements liés à ces émotions sont considérés comme faisant partie du répertoire comportemental de l'espèce humaine ? Quels sont les facteurs à l'origine de ces préférences ? Sont-ils réellement justifiés, ou bien dépendent-ils d'un effet de mode, de subjectivité ou de la dynamique socioculturelle d'une époque ?
Par ailleurs, l'étude du comportement basée principalement sur l'observation d'un sujet met en exergue la problématique béhavioriste de la dissociation entre les réactions motrices externes et les processus neurophysiologiques internes. Cette dissociation ne semble guère conforme à la réalité neurologique, où comportements et processus cérébraux relèvent d'une même unité fonctionnelle dynamique. Cette dissociation interne/externe est considéré dans le cadre de cette recherche plus comme un moyen didactique d'analyse et de description que comme une réalité psychobiologique.
Un autre problème serait la pertinence scientifique de l'étude d'un comportement qui apparemment semble bien caractérisé et défini, tels les comportements sexuel, maternel ou d'agression, mais qui en fait correspondraient au regroupement d'un ensemble d'actions motrices diverses, sous-tendu par des processus psychobiologiques différents. Exprimé autrement, qu'est-ce qui permet d'être certain que le comportement sexuel ou maternel existe au niveau neurobiologique ? Quels seraient alors les problèmes et les biais introduits par l'étude d'un objet conceptuel qui n'existe que dans l'esprit du chercheur ?
Proposition de définition
La première étape de l'élaboration d'une définition est d'identifier et de caractériser ce qui rend nécessaire la création de cette définition.
Une caractéristique majeure de certains organismes vivants est la capacité de mouvements, qui permet d'interagir activement avec l'environnement. Un aspect important de cette capacité est que le mouvement n'est pas un effet fortuit (comme le mouvement d'une branche au vent), mais le résultat d'une organisation spécifique de l'organisme.
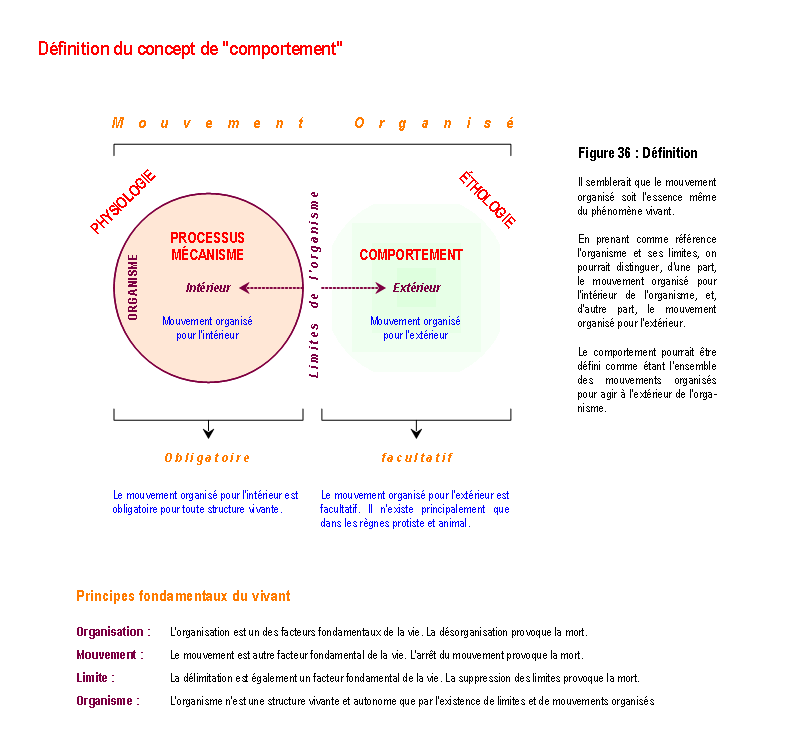
Les facteurs primordiaux du vivant qui sont directement en relation avec cette capacité de mouvement sont : l'organisation, le mouvement, la limite et l'organisme. L'organisation est un facteur primordial et fondamental du vivant : tout être vivant est une structure hautement organisée. La désorganisation provoque la mort. Le mouvement est un autre facteur primordial du vivant : quasiment tous les éléments d'une structure vivante sont en mouvement perpétuel. L'arrêt du mouvement provoque la mort. La limite est également un facteur primordial du vivant : tous les êtres vivants possèdent des limites externes et souvent de nombreuses limites internes. La suppression des limites provoque la mort. L'organisme n'est une structure vivante que par l'existence de limites et de mouvements organisés.
La synthèse de tous les éléments présentés ci-dessus permet d'élaborer une définition du concept de "comportement", basée sur les caractéristiques biologiques structurelles et fonctionnelles primordiales, et qui est valable pour tous les organismes vivants (Figure 36).
On observe que le mouvement organisé pour l'intérieur de l'organisme (réactions enzymatiques, transport intracellulaire des molécules, pompes protéiques, endocytose, migrations cellulaires, circulation sanguine, etc.) est une nécessité vitale absolue pour tous les êtres vivants. Par contre, le mouvement organisé pour l'extérieur de l'organisme (locomotion, préhension, etc.), qui correspond à la notion de "comportement", est facultatif. Il n'existe principalement que dans les règnes protiste et animal.
En fonction de cette synthèse, la définition du concept de "comportement", valable pour tous les organismes vivants, est : ensemble des mouvements organisés pour agir à l'extérieur de l'organisme.
Cette définition permet d'identifier sans ambiguïté tous les comportements : l'observation externe et objective d'un mouvement d'un organisme (ou d'une partie d'un organisme) est un comportement s'il existe des structures biologiques spécifiquement organisées pour produire ce mouvement ; par exemple le mouvement des yeux des mammifères n'est pas le résultat indirect des mouvements de la tête, mais résulte d'un contrôle neural (noyaux oculomoteurs III, IV et VI, etc.) des muscles oculaires : il existe donc un comportement oculaire.
En tenant compte de cette définition générale et des caractéristiques anatomo-fonctionnelles des mammifères, la définition proposée du concept de "comportement" pour l'Homme serait : Ensemble des mouvements produits par l'activation du système musculaire squelettique, sous contrôle de l'activité neurale motrice pyramidale et extrapyramidale.
Annexe II : Questionnaire
Enquête sur le comportement sexuel
Madame, Mademoiselle, Monsieur,
Nous sommes une équipe de chercheurs en sexologie et nous organisons, avec le concours de l'École Pratique des Hautes Études (Paris) et l'Observatoire International du Couple, une enquête sur le comportement sexuel.
Nous cherchons pour cette étude universitaire des personnes qui accepteraient de témoigner de leur expérience personnelle, en toute honnêteté et confidentialité, pour aider à mieux comprendre la sexualité.
Si vous souhaitez participer à cette étude, merci de bien vouloir répondre au questionnaire ci-joint, qui est entièrement anonyme. Si en complétant ce questionnaire vous changez d'avis, vous êtes libre de vous arrêter à tout moment.
Les données recueillies dans le cadre de cette recherche sont anonymes et resteront confidentielles. Vous pouvez à tout moment demander toute information complémentaire que vous souhaiteriez aux coordonnées indiquées ci-dessous.
Merci pour votre participation
Comment nous joindre ?
Dr. P. Brenot, Dr. C. Richard-Foy, Dr. S. Wunsch
49 bis avenue Jean-Jaurès, 33600 Pessac
Téléphone : 05 56 80 32 13 -- Fax : 05 56 45 10 14
Enquête sur le comportement sexuel
INTRODUCTION
Ce questionnaire s'adresse à un public très large et très divers. Certaines remarques ou précisions peuvent vous sembler inutiles ; des questions pourront vous paraître naïves, ou éventuellement choquantes, en fonction de vos connaissances ou de votre sensibilité. Nous vous remercions malgré tout de répondre à ce questionnaire consciencieusement et avec sincérité.
Merci donc de prendre un peu de temps (environ 30 mn) pour répondre à cette enquête qui est, bien entendu, entièrement anonyme.
Remarque importante : les réponses à certaines des questions ci-dessous se font par des notes. Toutes les notes vont de 0 à 9, et ce n'est pas un classement : vous pouvez donner plusieurs fois la même note, si nécessaire.
QUESTION 1 :
– Quelle est la ou les activités que vous préférez le plus ?
Donnez à toutes les activités ci-dessous une note allant de 0 (aucune préférence) à 9 (préférence maximale), en fonction de vos préférences.
Remarque : lisez d'abord les différentes rubriques, comparez, évaluez, puis notez.
…… Respirer des senteurs, des parfums, etc. (activité olfactive)
…… Manger ou goûter des aliments, des friandises, des boissons, etc. (activité gustative)
…… Faire l'amour avec mon partenaire (activité sexuelle)
…… Écouter de la musique, des chants, etc. (activité auditive)
…… Regarder un paysage, une œuvre d'art, des visages, etc. (activité visuelle)
…… Me masturber (activité sexuelle)
…… Lire une revue, écrire un poème, jouer aux échecs ou au Scrabble, etc. (activité intellectuelle)
…… Faire des activités physiques : baignade, sports, glisse, vitesse, vertige, etc. (activité corporelle)
…… Consommer de l'alcool, du tabac, une drogue, un médicament, ou vivre une situation non "naturelle" (activité "artificielle")
Autres activités que vous préférez le plus :
…… ……………………………………………………………………………………
…… ……………………………………………………………………………………
QUESTION 2 :
Nous appelons "coït vaginal" l'activité sexuelle qui consiste à introduire le pénis de l'homme dans le vagin de la femme. Nous appelons "coït anal" l'activité sexuelle qui consiste à introduire le pénis de l'homme dans l'anus de la femme ou de l'homme.
– Pouvez-vous vous rappeler à partir de quel âge vous avez eu les activités sexuelles listées ci-dessous ?
Remarque : Si vous ne vous rappelez pas de l'âge exact, donnez l'âge qui vous semble, en accord avec vos souvenirs, le plus vraisemblable. Mais si vous ne vous rappelez vraiment pas, ne mettez pas de fausse réponse. Mettez un point d'interrogation [ ? ]
– Avez-vous eu, au cours de votre vie, au moins une activité sexuelle avec vous-même (par exemple caresse de vos organes génitaux pour avoir du plaisir sexuel, ou masturbation pour ressentir l'orgasme, etc.)
o Oui, une ou quelques fois
o Oui, régulièrement
o Non
– Si OUI, votre âge lors de la première fois : ……… ans
– Si c'est régulier, à partir de quel âge la masturbation est-elle devenue répétée et habituelle : ……… ans
– Avez-vous eu, au cours de votre vie, au moins une activité sexuelle avec une autre personne (par exemple caresses érotiques des fesses ou des organes génitaux de l'autre personne, ou masturbation réciproque, ou baiser, ou fellation, ou coït vaginal ou anal, etc.) ?
o Oui, une ou quelques fois
o Oui, régulièrement
o Non
– Si OUI, votre âge lors de la première fois : ……… ans
– Si c'est régulier, à partir de quel âge ces activités sont-elles devenues répétées et habituelles : ……… ans
– Avez-vous eu, au cours de votre vie, au moins un coït vaginal (pénétration du pénis dans le vagin) avec une autre personne ?
o Oui, une ou quelques fois
o Oui, régulièrement
o Non
– Si OUI, votre âge lors du premier coït vaginal : ……… ans
– Si c'est régulier, à partir de quel âge le coït vaginal est-il devenu répété et habituel : ……… ans
– Avez-vous eu, au cours de votre vie, au moins un coït anal (actif ou passif) avec une autre personne ?
o Oui, une ou quelques fois
o Oui, régulièrement
o Non
– Si OUI, votre âge lors du premier coït anal : ……… ans
– Si c'est régulier, à partir de quel âge le coït anal est-il devenu répété et habituel : ……… ans
QUESTION 3 :
– Quand vous n'avez pas de partenaire, quelle est, en général, la fréquence de vos activités sexuelles avec vous-même ?
o Plusieurs fois par jour
o Une fois par jour
o Plusieurs fois par semaine
o Une fois par semaine
o Plusieurs fois par mois
o Une fois par mois
o Moins d'une fois par mois
o Durant toute ma vie, moins de cinq activités sexuelles
o Aucune activité sexuelle
QUESTION 4 :
– Quand vous êtes seul et que vous vous masturbez, quelle est la ou les sources de votre excitation sexuelle ?
a) Cochez la case des sources d'excitation sexuelle que vous utilisez quand vous vous masturbez.
b) Puis donnez à chacune de vos sources d'excitation une note allant de 0 (aucune excitation) à 9 (excitation maximale), en fonction de son importance pour provoquer l'excitation sexuelle.
Remarque : Comparez d'abord vos différentes sources, évaluez-les, puis notez.
…… o Des images érotiques fixes (photographies, dessins, etc.)
…… o Des images érotiques animées (films, vidéos, etc.)
…… o Des lectures érotiques
…… o Des discussions érotiques par Internet / Minitel / Téléphone
…… o Des souvenirs d'activités sexuelles que vous avez vues (souvenirs de scènes sexuelles)
…… o Des souvenirs de vos activités sexuelles avec vos partenaires (souvenirs de vécus sexuels)
…… o Des situations et des activités sexuelles que vous imaginez (fantasmes sexuels)
…… o Une combinaison de plusieurs sources : par exemple, des souvenirs avec des fantasmes tout en regardant en même temps des images ou une vidéo
…… o En observant (discrètement) une autre personne
…… o Uniquement la stimulation corporelle, sans aucune discussion, image, pensée ou souvenir
…… o Autres sources : ……………………………………………………
o Je ne me masturbe pas, ou je ne me suis pas assez masturbé pour pouvoir répondre à cette question
ATTENTION : Si vous n'avez jamais eu d'activités sexuelles avec un partenaire (absolument rien, même pas un ou quelques baisers ou quelques caresses érotiques), allez à la question n°11
QUESTION 5 :
– Avez-vous actuellement des activités sexuelles avec un (ou plusieurs) partenaire(s) ?
o Oui o Non
1) Si vous avez répondu NON, depuis combien de temps
n'avez-vous plus eu d'activités sexuelles avec un partenaire : ……… ans [ ou ……… mois ]
2) Si vous avez répondu OUI, êtes-vous actuellement amoureux(se) de votre partenaire ?
o Oui o Non
QUESTION 6 :
– Quand vous êtes avec un partenaire (ou que des partenaires sont disponibles), quelle est, en général, la fréquence de vos activités sexuelles avec ce (ou ces) partenaire(s) ?
o Plusieurs fois par jour
o Une fois par jour
o Plusieurs fois par semaine
o Une fois par semaine
o Plusieurs fois par mois
o Une fois par mois
o Moins d'une fois par mois
o Durant toute ma vie, moins de cinq activités sexuelles
o Aucune activité sexuelle
QUESTION 7 :
– Durant toute votre vie, quel a été le nombre de vos partenaires sexuels ?
o Plus de 50 partenaires
o Entre 21 et 50 partenaires
o Entre 11 et 20 partenaires
o Entre 6 et 10 partenaires
o Entre 3 et 5 partenaires
o 2 partenaires
o 1 partenaire
o Aucun
QUESTION 8 :
– En prenant en compte toutes vos activités sexuelles avec d'autre(s) personne(s), depuis vos premières activités jusqu'à aujourd'hui, vos activités sexuelles sont-elles plutôt :
o Exclusivement homosexuelles
o Majoritairement homosexuelles
o Bisexuelles, mais plutôt homosexuelles
o Bisexuelles, autant hétérosexuelles qu'homosexuelles
o Bisexuelles, mais plutôt hétérosexuelles
o Majoritairement hétérosexuelles
o Exclusivement hétérosexuelles
Remarque : Avoir des activités homosexuelles ne signifie pas qu'on est "homosexuel".
QUESTION 9 :
– Au cours de votre vie, avez-vous pratiqué les activités suivantes ?
– L'échangisme
o Non
o Au moins une fois
o Plusieurs fois
o Régulièrement (au moins plusieurs fois par an)
– Le triolisme (vous et deux partenaires)
o Non
o Au moins une fois
o Plusieurs fois
o Régulièrement (au moins plusieurs fois par an)
– La sexualité en groupe (vous et plus de deux partenaires)
o Non
o Au moins une fois
o Plusieurs fois
o Régulièrement (au moins plusieurs fois par an)
– Autres activités sexuelles pratiquées
(Ondinisme, SM, Bondage, etc.) : ………………………………………
o Non Ondinisme = activités sexuelles avec de l'urine
o Au moins une fois SM = activités sexuelles sado-masochistes
o Plusieurs fois Bondage = un des partenaires sexuels est attaché
o Régulièrement (au moins plusieurs fois par an)
QUESTION 10 :
– Donnez la ou les principales raisons de vos activités sexuelles avec un partenaire.
a) Cochez la case de la ou des raisons qui font que vous avez des activités sexuelles avec votre partenaire.
(Attention de bien séparer les raisons qui font que vous avez envie d'être avec un partenaire, des raisons pour lesquelles vous avez envie d'avoir des activités sexuelles avec ce partenaire)
b) Puis donnez à chacune de vos raisons une note allant de 0 (aucune importance) à 9 (importance principale), en fonction de son importance pour expliquer vos activités sexuelles avec votre partenaire.
Remarque : Comparez d'abord vos différentes raisons, évaluez-les, puis notez.
…… o C'est mon (ou ma) partenaire qui me les demande
…… o Pour se faire des papouilles, des chatouilles, des bisous ou des câlins coquins et malins
…… o Pour se donner du plaisir
…… o Pour le sentiment, dans les contacts intimes, d'être accepté et aimé
…… o Parce que j'aime mon (ou ma) partenaire
…… o Pour recevoir de la tendresse physique
…… o Pour le contact physique intime contre la peau et le corps de mon partenaire
…… o Pour la complicité qui existe entre nous durant les activités sexuelles
…… o Pour le sentiment, dans les contacts intimes, d'être en sécurité
…… o Pour chahuter ou lutter dans des corps à corps érotiques et passionnés
…… o Parce que je ressens comme une tendance ou un besoin qui me pousse à faire des activités sexuelles
…… o Pour le désir d'être pénétrée (ou de pénétrer) mon (ou ma) partenaire
…… o Pour avoir des enfants
…… o C'est ce qui est normal de faire avec un partenaire
…… o La curiosité
…… o Autre raison : …………………………………………………………
o Je n'arrive pas à identifier la ou les raisons
QUESTION 11 :
– Si vous étiez totalement libre, et qu'il n'existait aucun problème pour rencontrer (ou ne pas rencontrer) des partenaires, quelle serait la fréquence de vos activités sexuelles avec des partenaires ?
o Plusieurs fois par jour
o Une fois par jour
o Plusieurs fois par semaine
o Une fois par semaine
o Plusieurs fois par mois
o Une fois par mois
o Moins d'une fois par mois
o Aucune activité sexuelle
QUESTION 12 :
Nous appelons "plaisir sexuel" toutes les différentes sensations très agréables et particulières, voluptueuses, qui sont provoquées par les caresses et les stimulations, entre autres, du pénis chez l'homme et du clitoris chez la femme. Nous appelons "orgasme" le plaisir sexuel le plus intense, qui se déclenche subitement et se traduit par un maximum de plaisir, bref et parfois extatique.
– Pouvez-vous vous rappeler à partir de quel âge vous avez ressenti les différents plaisirs listés ci-dessous ?
Remarque : Si vous ne vous rappelez pas de l'âge exact, donnez l'âge qui vous semble, en accord avec vos souvenirs, le plus vraisemblable. Mais si vous ne vous rappelez vraiment pas, ne mettez pas de fausse réponse. Mettez un point d'interrogation [ ? ]
– Avez-vous déjà ressenti du plaisir, autre que du plaisir sexuel (par exemple le plaisir d'un bon repas, le plaisir d'écouter de la musique, etc.) ?
o Oui o Non
Si OUI, votre âge lors de la première sensation de plaisir
non sexuel dont vous pouvez vous souvenir : ……… ans
– Avez-vous déjà ressenti du plaisir sexuel, autre que l'orgasme (par exemple le plaisir sexuel d'être caressé sur les fesses, ou le plaisir sexuel d'embrasser, ou de se caresser sensuellement, etc.) ?
o Oui, une ou quelques fois
o Oui, régulièrement
o Non
– Si OUI, votre âge lors du premier plaisir sexuel dont vous pouvez vous souvenir : ……… ans
– Si c'est régulier, à partir de quel âge le plaisir sexuel est-il devenu répété et habituel : ……… ans
– Avez-vous déjà ressenti un orgasme ?
o Oui, une ou quelques fois
o Oui, régulièrement
o Non
– Si OUI, votre âge lors du premier orgasme dont vous pouvez vous souvenir : ……… ans
– Si c'est régulier, à partir de quel âge l'orgasme est-il devenu répété et habituel : ……… ans
QUESTION 13 :
– Pouvez-vous comparer les différents types de plaisir que vous avez déjà vécus ?
Donnez à tous les plaisirs ci-dessous une note allant de 0 (absence de plaisir) à 9 (plaisir extrême, extase), en fonction de ce que vous avez vécu et ressenti.
Remarque : lisez d'abord les différentes rubriques, comparez, évaluez, puis notez.
…… Le plaisir sexuel (incluant l'orgasme) le plus intense que j’ai vécu, provoqué par moi-même (masturbation, etc.)
…… Le plaisir sexuel (incluant l'orgasme) le plus intense que j’ai vécu, provoqué par un partenaire
…… Le plaisir émotionnel (bien-être, gaieté, joie, bonheur, etc.) le plus intense que j'ai vécu
…… Le plaisir gustatif (aliments, boissons, friandises, etc.) le plus intense que j'ai vécu
…… Le plaisir olfactif (odeurs, parfums, senteurs, etc.) le plus intense que j'ai vécu
…… Le plaisir auditif (sons, musique, chants, etc.) le plus intense que j'ai vécu
…… Le plaisir visuel (visages, paysages, œuvre d'art, etc.) le plus intense que j'ai vécu
…… Le plaisir intellectuel (lecture, humour, jouer aux échecs ou au Scrabble, etc.) le plus intense que j'ai vécu
…… Le plaisir corporel (baignade, activité physique, sports de glisse, de vitesse ou de vertige, etc.) le plus intense que j'ai vécu
…… Le plaisir artificiel (drogue, alcool, tabac, médicament, ou autre situation non ‘naturelle’) le plus intense que j'ai vécu
QUESTION 14 :
– Quelle est la période de votre vie où vous avez vécu les plaisirs sexuels les plus intenses ?
Remarque : Si vous avez vécu plusieurs périodes ayant des pics de plaisir sexuel, prenez la période où le plaisir était le plus intense.
– À quel âge à débuté cette période : ….…… ans
– Quelle est la durée de cette période : ……… ans [ ou ….…… mois ]
– Durant cette période, quelle était la fréquence de vos plaisirs sexuels ?
o Plusieurs fois par jour
o Une fois par jour
o Plusieurs fois par semaine
o Une fois par semaine
o Plusieurs fois par mois
o Une fois par mois
o Moins d'une fois par mois
o Durant toute ma vie, moins de cinq plaisirs sexuels
QUESTION 15 :
– Quelle est, pour vous, l'importance du plaisir sexuel dans votre vie ?
o Très important
o Important
o Moyennement important
o Peu important
o Sans importance
– Quand vous avez des activités sexuelles, atteindre l'orgasme est-il important pour vous ?
o Très important
o Important
o Moyennement important
o Peu important
o Sans importance
o Je ne peux pas répondre à cette question, je n'ai jamais ressenti d'orgasme
QUESTION 16 : ATTENTION : uniquement pour les FEMMES
– Pouvez-vous comparer l'intensité du plaisir sexuel provoqué par la stimulation des différentes parties du corps ?
a) Que la stimulation soit faite par vous-même ou par un partenaire, donnez, à toutes les régions de votre corps qui sont stimulées durant les activités sexuelles, une note allant de 0 à 9, en fonction du plaisir sexuel le plus intense qu'elle vous a procuré.
Remarque : lisez d'abord les différentes rubriques, comparez, évaluez, puis notez. ( 0 = absence de plaisir, 9 = plaisir extrême, extase)
b) Cochez la case "Orgasme" si la stimulation uniquement de cette région peut, chez vous, provoquer un orgasme.
Tête note de 0 à 9 : ………… o Orgasme
Lèvres note de 0 à 9 : ………… o Orgasme
Langue note de 0 à 9 : ………… o Orgasme
Seins note de 0 à 9 : ………… o Orgasme
Ventre note de 0 à 9 : ………… o Orgasme
Clitoris note de 0 à 9 : ………… o Orgasme
Vagin note de 0 à 9 : ………… o Orgasme
Dos note de 0 à 9 : ………… o Orgasme
Fesses note de 0 à 9 : ………… o Orgasme
Anus note de 0 à 9 : ………… o Orgasme
Une zone très sensible à l'intérieur du vagin (Point G), que votre partenaire stimule avec ses doigts
note de 0 à 9 : ………… o Orgasme
Autres régions :
…………..…………………… note de 0 à 9 : ………… o Orgasme
…………..…………………… note de 0 à 9 : ………… o Orgasme
QUESTION 17 : ATTENTION : uniquement pour les FEMMES
Si vous n'avez jamais eu d'activités sexuelles avec un partenaire, allez à la question n°20
– Quelle est, parmi toutes les activités sexuelles que vous avez déjà vécues, la ou les activités sexuelles que vous préférez faire avec votre (ou vos) partenaire(s) ?
a) Cochez la case des activités sexuelles que vous avez déjà pratiquées.
b) Puis donnez à chacune de vos activités sexuelles une note allant 0 (aucune préférence) à 9 (préférence maximale), en fonction de vos préférences.
Remarque : Comparez d'abord vos différentes activités sexuelles, évaluez-les, puis notez.
…… o Embrasser mon (ma) partenaire avec la langue
…… o Faire une fellation à mon partenaire Fellation = stimuler et sucer le pénis avec la bouche
…… o Faire un cunnilingus à ma partenaire Cunnilingus = stimuler le clitoris avec la langue
…… o Mon (ma) partenaire me fait un cunnilingus
…… o Être en contact physique intime : se serrer dans les bras, être l'un sur l'autre, etc.
…… o Masturber mon (ma) partenaire
…… o Être masturbé par mon (ma) partenaire
…… o Se masturber réciproquement
…… o Faire des stimulations érotiques des fesses / anus / rectum de mon (ma) partenaire avec les mains
…… o Mon (ma) partenaire me fait des stimulations érotiques des fesses / anus / rectum avec les mains
…… o Se faire des caresses sensuelles
…… o Mon partenaire me fait un coït anal
…… o Mon partenaire me fait un coït vaginal
…… o Faire un 69 : fellation / cunnilingus réciproque
Autres activités sexuelles que vous préférez le plus :
…… ……………………………………………………………………………………
…… ……………………………………………………………………………………
QUESTION 16 : ATTENTION : uniquement pour les HOMMES
– Pouvez-vous comparer l'intensité du plaisir sexuel provoqué par la stimulation des différentes parties du corps ?
a) Que la stimulation soit faite par vous-même ou par un partenaire, donnez, à toutes les régions de votre corps qui sont stimulées durant les activités sexuelles, une note allant de 0 à 9, en fonction du plaisir sexuel le plus intense qu'elle vous a procuré.
Remarque : lisez d'abord les différentes rubriques, comparez, évaluez, puis notez. ( 0 = absence de plaisir, 9 = plaisir extrême, extase)
b) Cochez la case "Orgasme" si la stimulation uniquement de cette région peut, chez vous, provoquer un orgasme.
Tête note de 0 à 9 : ………… o Orgasme
Lèvres note de 0 à 9 : ………… o Orgasme
Langue note de 0 à 9 : ………… o Orgasme
Poitrine note de 0 à 9 : ………… o Orgasme
Ventre note de 0 à 9 : ………… o Orgasme
Pénis note de 0 à 9 : ………… o Orgasme
Testicule note de 0 à 9 : ………… o Orgasme
Dos note de 0 à 9 : ………… o Orgasme
Fesses note de 0 à 9 : ………… o Orgasme
Anus note de 0 à 9 : ………… o Orgasme
Une zone très sensible à l'intérieur de l'anus (Prostate), que votre partenaire stimule avec ses doigts
note de 0 à 9 : ………… o Orgasme
Autres régions :
…………..…………………… note de 0 à 9 : ………… o Orgasme
…………..…………………… note de 0 à 9 : ………… o Orgasme
QUESTION 17 : ATTENTION : uniquement pour les HOMMES
Si vous n'avez jamais eu d'activités sexuelles avec un partenaire, allez à la question n°20
– Quelle est, parmi toutes les activités sexuelles que vous avez déjà vécues, la ou les activités sexuelles que vous préférez faire avec votre (ou vos) partenaire(s) ?
a) Cochez la case des activités sexuelles que vous avez déjà pratiquées.
b) Puis donnez à chacune de vos activités sexuelles une note allant 0 (aucune préférence) à 9 (préférence maximale), en fonction de vos préférences.
Remarque : Comparez d'abord vos différentes activités sexuelles, évaluez-les, puis notez.
…… o Embrasser ma (mon) partenaire avec la langue
…… o Faire une fellation à mon partenaire Fellation = stimuler et sucer le pénis avec la bouche
…… o Faire un cunnilingus à ma partenaire Cunnilingus = stimuler le clitoris avec la langue
…… o Ma (mon) partenaire me fait une fellation
…… o Être en contact physique intime : se serrer dans les bras, être l'un sur l'autre, etc.
…… o Masturber ma (mon) partenaire
…… o Être masturbé par ma (mon) partenaire
…… o Se masturber réciproquement
…… o Faire des stimulations érotiques des fesses / anus / rectum de ma (mon) partenaire avec les mains
…… o Ma (mon) partenaire me fait des stimulations érotiques des fesses / anus / rectum avec les mains
…… o Se faire des caresses sensuelles
…… o Mon partenaire me fait un coït anal
…… o Faire un coït vaginal à ma partenaire
…… o Faire un coït anal à ma (mon) partenaire
…… o Faire un 69 : fellation / cunnilingus réciproque
Autres activités sexuelles que vous préférez le plus :
…… ……………………………………………………………………………………
…… ……………………………………………………………………………………
QUESTION 18 :
– Préférez-vous les sensations de plaisir sexuel :
o Provoquées par vous-même (masturbation, etc.).
o Provoquées par votre partenaire (activités sexuelles en couple).
o Provoquées avec votre partenaire à stimulations réciproques en même temps.
o Provoquées par plusieurs partenaires (activités sexuelles en groupe).
o Je n'ai pas de préférence.
QUESTION 19 :
– Préférez-vous avoir des activités sexuelles :
o Seul(e) (masturbation, etc.).
o En couple, avec votre partenaire.
o En groupe, avec plusieurs partenaires.
o Je n'ai pas de préférence.
QUESTION 20 :
– Avez-vous des fantasmes sexuels ?
o Très souvent
o Souvent
o Rarement
o Jamais
– En général, comment surviennent-ils ?
o Involontairement, de manière spontanée
o Volontairement, quand j'en ai envie
o Volontairement et involontairement
– Quel est votre fantasme le plus stimulant ?
o Une scène érotique avec votre partenaire
o Une scène érotique avec un(e) autre partenaire
o Une scène romantique
o Une sexualité de groupe (orgie, échangisme, triolisme)
o Autre fantasme
– Dans vos fantasmes, les activités sexuelles sont plutôt :
o Homosexuelles
o Bisexuelles
o Hétérosexuelles
QUESTION 21 :
– En général, quelle est l'intensité de votre désir d'avoir des activités sexuelles avec un partenaire ?
Remarque : Entourez le nombre qui représente le mieux la force de votre désir sexuel.
0 1 2 3 4 5 6 7 8 9
absence désir fort,
de désir intense
QUESTION 22 :
– Quelle est, pour vous, l'importance de la sexualité dans votre vie ?
o N'a aucune importance.
o A une importance mineure.
o Est d'une importance secondaire.
o A une importance égale à celle des autres aspects de la vie.
o Est relativement important.
o A une grande importance.
o Est un aspect central de mon existence.
QUESTION 23 :
– Ressentez-vous de la honte, ou de la pudeur, ou de la gêne en situation d’intimité avec votre partenaire ?
o Beaucoup.
o Moyennement.
o Un peu.
o Pas du tout.
QUESTION 24 :
1) Avez-vous vécu, dans l’enfance, l’adolescence ou l'âge adulte, un ou plusieurs actes sexuels qui vous ont provoqué des émotions négatives intenses (peur, douleur, dégoût, honte, culpabilité …) ?
o Oui o Non
Si vous avez répondu OUI, pouvez-vous indiquer quelles sont cette ou ces expériences sexuelles négatives ?
………………………………………………………………………………………………………………………………………………………………………
………………………………………………………………………………………………………………………………………………………………………
Cette ou ces expériences sexuelles négatives sont-elles une gêne ou un handicap pour votre sexualité actuelle ?
o Oui o Non
2) Avez-vous un facteur psychologique qui perturbe actuellement votre sexualité (timidité importante, peur des MST, agressivité, dépression, toxicomanie, ou éducation anti-sexuelle, etc.) ?
o Oui o Non
Si vous avez répondu OUI, pouvez-vous indiquer quel est ce (ou ces) facteurs ?
………………………………………………………………………………………………………………………………………………………………………
………………………………………………………………………………………………………………………………………………………………………
QUESTION 25 :
– Avez-vous actuellement un problème physiologique ou médical qui perturbe votre sexualité (douleur au cours des rapports, trouble de l'érection, vaginisme, éjaculation précoce, obésité, handicap moteur, etc.) ?
o Oui o Non
Si vous avez répondu OUI, pouvez-vous indiquer lequel (ou lesquels) ?
………………………………………………………………………………………………………………………………………………………………………
………………………………………………………………………………………………………………………………………………………………………
QUESTION 26 :
– Avez-vous ressenti de la gêne ou étiez-vous mal à l'aise en répondant à ces questions ?
o Beaucoup de gêne / Très mal à l'aise.
o De la gêne / Mal à l'aise.
o Un peu de gêne / Un peu mal à l'aise.
o Aucune gêne, mais parler de ma sexualité est inhabituel.
o Aucune gêne, j'ai l'habitude de parler de ma sexualité.
Renseignements
Pouvez-vous mentionner votre :
– Âge :
– Sexe :
– Votre milieu d'origine :
o Rural
o Ville < 10.000 habitants
o Ville > 10.000 habitants
Vous avez terminé le questionnaire. Merci de votre participation à cette recherche.
Annexe III : Résultats et analyses de l'enquête par questionnaire
1 – Annexe du chapitre "Matériel & Méthode"
a – Critères de sélection des participants & Formations des groupes
a – Critères de sélection
Comme l'objectif principal de cette enquête est l'analyse du rôle et de l'importance du plaisir érotique dans la sexualité, il est très important que les sujets sélectionnés pour être étudiés n'aient pas de problèmes qui perturbent soit leurs perceptions psychiques des sensations érotiques, soit leur comportement sexuel.
Il est bien sûr évident que tous les facteurs d'influence négative ne pourront être ni connus ni écartés. Le contexte social et culturel a une influence majeure sur le développement de la sexualité, et il n'est pas possible dans le cadre de cette thèse de mener des études expérimentales psychologiques, sociologiques et transculturelles permettant d'évaluer puis de contrôler toutes les caractéristiques et l'impact de tous les facteurs psychiques et socioculturels. Par contre, il est possible d'écarter une partie des facteurs à l'origine de divers types de troubles, en particulier ceux qui sont les plus facilement identifiables. La dernière section du questionnaire a été spécialement conçue afin de recueillir des informations permettant d'identifier les troubles les plus perceptibles. Par ailleurs, l'analyse des réponses et des éventuels commentaires des participants, tant sur la forme que sur le fond, donne également des informations qui permettent d'identifier et d'évaluer d'éventuels problèmes.
En fonction des éléments présentés ci-dessus, des critères de sélection, d'exclusion et d'analyse séparée ont été sélectionnés.
Présentation des critères
Critère de sélection
Les critères de sélections permettent de décider, toujours à partir des mêmes critères, quels questionnaires seront retenus pour tester les hypothèses.
Les critères de sélection sont :
– Questionnaire complètement et correctement remplis.
– Absence de troubles physiologiques ou psychologiques perturbant la sexualité.
– Personnes majeures.
Critères d'exclusion
Les critères d'exclusion permettent de décider quels questionnaires seront complètement exclu de l'étude.
Les critères d'exclusion sont :
– Questionnaire incomplet.
– Questionnaire avec trop d'erreurs.
– Questionnaire complété de manière fantaisiste ou incohérente.
Critère d'analyse séparée
Les critères d'analyse séparée permettent de décider quels questionnaires seront analysés à part, afin d'obtenir des informations complémentaires.
Les critères d'analyse séparée sont :
– Existence de troubles physiologiques perturbant la sexualité.
– Existence de troubles psychologiques perturbant la sexualité.
Explicitation des critères
Explication du critère "Questionnaire incomplet"
Le critère de complétude n'est pas aussi simple qu'il paraît au premier abord. En effet, il est possible de trouver un questionnaire où ne manque que quelques items, en raison d'une mauvaise compréhension d'une consigne, d'une inattention, ou d'un autre facteur indépendant d'une volonté de bien faire. Néanmoins, tant que ces quelques omissions n'altèrent pas la qualité des autres réponses données, le questionnaire est exploitable.
Le questionnaire incomplet qu'il faut absolument exclure est celui où il devient évident que la personne n'a pas répondu à certaines questions parce qu'elle n'était pas ou plus dans un état d'implication et de motivation à répondre sérieusement et consciencieusement. Dans ce cas, les réponses déjà données ne sont vraisemblablement pas fiables et doivent être exclues de l'étude.
Au final, la procédure retenue est la suivante : un questionnaire incomplet qui n'a pas été rempli jusqu'à la fin est exclu. Un questionnaire incomplet remplit jusqu'à la fin, mais où il manque toutes les réponses à au moins une des questions à multiples réponses (n°1, 2, 4, etc.) et exclu si la personne a indiqué qu'elle trouvait le questionnaire trop long. Enfin, dans les autres cas, le questionnaire incomplet est considéré comme contenant des erreurs, et est traité en fonction des critères spécifiques à l'analyse des erreurs (voir ci-dessous).
Explication du critère "Questionnaire avec trop d'erreurs"
De nombreux facteurs peuvent être à l'origine d'erreurs dans les réponses au questionnaire (inattention, mauvaise compréhension d'une consigne, erreur de frappe, etc.). L'exclusion systématique, par précaution, de tous les questionnaire ayant ne serait-ce qu'une erreur conduit à supprimer un grand nombre de données qui pourraient être exploitables.
Afin de détecter certains types d'erreurs, le questionnaire a été élaboré avec des questions redondantes qui permettent, par recoupement, d'analyser la cohérence des réponses et de vérifier si la personne a répondue de manière sincère, fantaisiste ou si elle n'a pas un niveau de compréhension suffisant.
Au final, la procédure retenue est la suivante : toutes les erreurs sont comptabilisées et le questionnaire est exclu s'il contient plus de 3 erreurs. Au delà de ce seuil, on peut considérer qu'il existe un problème qui affecte vraisemblablement toutes les réponses. En deçà de ce seuil, les erreurs relevées sont soit corrigées s'il est possible de comprendre l'erreur et de trouver la réponse correcte, soit la réponse erronée est supprimée et les autres données sont conservées pour analyses.
Explication du critère "Troubles physiologiques"
Les personnes ayant des handicaps ou des problèmes physiologiques qui perturbent leur sexualité sont analysées à part. Ces informations sont recueillies grâce à la question n° 25, et dépendent donc de l'honnêteté des participants et de leur capacité à identifier ces problèmes.
Comme exemple de troubles physiologiques, on peut donner : l'anhédonie, l'anorgasmie, ou spécifiquement pour les femmes l'hirsutisme, le vaginisme, les conséquences physiques de violences, les douleurs durant le coït, etc. ; pour les hommes les dysfonctions érectiles, l'éjaculation précoce, les douleurs péniennes, etc.
Les personnes souffrant de troubles bénins, occasionnels ou sans impact spécifique sur la sexualité ne sont pas exclues de l'enquête. On peut donner comme exemple la grossesse, qui est un état occasionnel et qui surtout est perçu comme absolument "normal", c'est-à-dire qu'il ne risque pas d'entraîner des problématiques psychologiques connexes négatives qui vont modifier les réactions des personnes.
Au final, la procédure retenue est la suivante : toute personne indiquant qu'elle a actuellement un problème physiologique qui perturbe sa sexualité est analysée à part, sauf si le problème, quand il est indiqué, est mineur et ne peut induire des modifications aux réponses du questionnaire.
Explication du critère "Troubles psychologiques"
Les personnes ayant des problèmes psychologiques qui perturbent leur sexualité sont analysées à part. Ces informations sont recueillies par la question n° 24, ainsi que les questions n° 23 et 26. Ces informations dépendent donc de l'honnêteté des participants et de leur capacité à identifier ces problèmes.
Seules les personnes donnant suffisamment de détails et de précisions permettant d'évaluer le trouble psychique comme mineur sont, après justifications, conservées.
Au final, la procédure retenue est la suivante : toute personne indiquant qu'elle a actuellement un problème psychologique qui perturbe sa sexualité est analysée à part, sauf si le problème, quand il est indiqué, est mineur et ne peut induire des modifications aux réponses du questionnaire. Les personnes indiquant ressentir moyennement ou beaucoup de gène aux questions 23 et 26 sont également analysées à part. Enfin, les personnes indiquant à la question n°12 ne pas ressentir un des types de plaisir (non sexuel, sexuel ou orgastique) sont elles aussi analysées à part.
b – Formations des groupes
Critères de formation des groupes
Les participants à cette étude qui ont été sélectionnés en fonction des critères ci-dessus sont ensuite répartit dans différents groupes, afin de tester les hypothèses relatives au rôle et à l'importance du plaisir érotique. Ces groupes sont créés en fonction de facteurs directement lié au plaisir érotique et aux activités érotiques. Ces facteurs, a priori, font que les personnes sont intrinsèquement différentes des autres ou que certaines caractéristiques les ont rendu différentes des autres. Ces facteurs sont :
– Le genre.
Les différences anatomiques et physiologiques entre les femmes et les hommes devraient entraîner des différences dans la perception de certaines sensations érotiques et, surtout, des différences dans les préférences pour certaines activités érotiques.
– L'orientation des activités érotiques.
Des pratiques érotiques très différentes sont susceptibles de modifier certaines sensations érotiques et la préférence pour certaines activités érotiques.
– L'absence ou la présence de l'orgasme.
L'orgasme étant considéré comme un des plaisirs le plus intenses que l'organisme peut éprouver, sont absence, qui en général provient de cas particuliers, doit vraisemblablement modifier l'évaluation du plaisir érotique et peut-être modifier certaines pratiques érotiques.
– L'absence ou la présence de masturbation.
L'absence de masturbation traduit en général soit un problème psychologique ou physiologique, soit des valeurs culturelles particulières. Toutes ces raisons justifient une analyse séparée.
– L'absence ou la présence d'activités érotiques.
Les personnes n'ayant pas d'activités érotiques avec des partenaires, ou n'ayant aucune activité érotique, même autoérotique, ont par rapport à la sexualité un vécu très différent des autres personnes, ce qui nécessite une analyse à part.
Les différents groupes ainsi constitués sont présentés dans le tableau ci-contre (Tableau 33).
b – Caractéristiques des participants à l'étude
Les pages suivantes contiennent une description du profil des activités érotiques des 5 principaux groupes (femmes hétérosexuelles et bisexuelles ; hommes hétérosexuels, bisexuels et homosexuels), puis une analyse de leurs désirs et fantasmes érotiques. Ces données sont utilisées afin d'obtenir des informations complémentaires et permettent également de savoir dans quelle mesure il sera possible, au terme de toutes les analyses, de regrouper ces différents groupes initiaux.
( NB : dans tous les tableaux de cette section, les nombres en italique correspondent aux personnes les plus bisexuelles, 3, 4 et 5 sur l'échelle de Kinsey )
b.1 – Profil des activités érotiques
Cette section contient la description du profil des activités érotiques des groupes sélectionnés pour tester l'hypothèse Hy .
a – Période du développement des activités érotiques
Les tableaux et figures ci-dessous indiquent les périodes d'initiation aux différentes pratiques érotiques, en fonction du genre et de l'orientation des activités sexuelles. Pour chaque type d'activités érotiques, les sujets ont donné l'âge de la première activité et l'âge à partir duquel cette activité est devenue régulière.
( réponses à la question n° 2 ; NB (rappel) : les nombres en italique correspondent aux personnes les plus bisexuelles, 3, 4 et 5 sur l'échelle de Kinsey )
|
Groupe (genre + orientation) |
Age première masturbation |
Age masturbation régulier |
Age premier sexe avec partenaire |
Age sexe avec partenaire régulier |
Age premier coit vaginal |
Age coit vaginal régulier |
Age premier coit anal |
Age coit anal régulier |
|
|
H_Hété |
Mean |
13,20 |
15,38 |
17,12 |
19,47 |
18,63 |
20,12 |
25,45 |
24,75 |
|
N |
113 |
76 |
112 |
77 |
108 |
76 |
67 |
16 |
|
|
Std. Deviation |
4,272 |
6,709 |
4,078 |
3,390 |
3,362 |
2,989 |
9,373 |
9,883 |
|
|
F_Hété |
Mean |
13,97 |
17,60 |
16,56 |
18,17 |
17,64 |
18,39 |
21,75 |
26,30 |
|
N |
129 |
66 |
132 |
76 |
132 |
74 |
93 |
20 |
|
|
Std. Deviation |
5,381 |
6,658 |
3,146 |
3,704 |
3,068 |
3,526 |
4,984 |
9,609 |
|
|
F_Bi |
Mean |
12,13 11,53 |
15,36 14,63 |
14,34 13,42 |
16,66 16,47 |
16,31 16,32 |
17,10 16,80 |
20,93 20,22 |
23,08 21,50 |
|
N |
56 |
28 |
58 |
32 |
57 |
29 |
46 |
12 |
|
|
Std. Deviation |
5,580 |
6,243 |
3,406 |
2,336 |
2,524 |
2,076 |
5,276 |
7,716 |
|
|
H_Bi |
Mean |
11,42 10,86 |
12,66 11,82 |
15,27 14,43 |
17,50 16,63 |
18,07 17,57 |
19,97 19,50 |
22,72 20,65 |
22,23 17,00 |
|
N |
62 |
38 |
59 |
38 |
60 |
33 |
53 |
13 |
|
|
Std. Deviation |
2,229 |
2,474 |
3,295 |
3,930 |
3,588 |
4,870 |
9,037 |
8,228 |
|
|
H_Homo |
Mean |
10,93 |
13,00 |
17,00 |
19,50 |
|
|
20,46 |
20,00 |
|
N |
15 |
4 |
16 |
2 |
|
|
13 |
2 |
|
|
Std. Deviation |
2,789 |
1,414 |
4,676 |
2,121 |
|
|
3,886 |
,000 |
|
|
Total |
Mean |
12,92 |
15,54 |
16,20 |
18,30 |
17,80 |
19,08 |
22,65 |
24,25 |
|
N |
375 |
212 |
377 |
225 |
357 |
212 |
272 |
63 |
|
|
Std. Deviation |
4,699 |
6,221 |
3,695 |
3,578 |
3,253 |
3,586 |
7,331 |
8,872 |
|
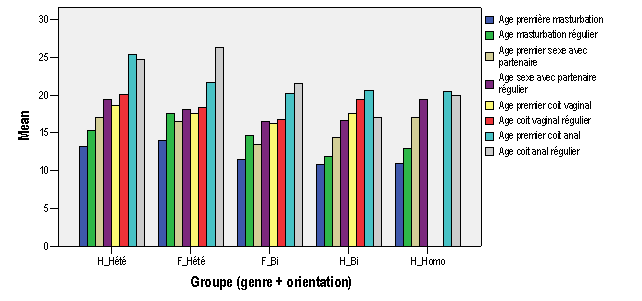
On observe pour tous les groupes que l'entrée dans la sexualité est progressive et qu'elle débute en général par la masturbation, en moyenne entre l'âge de 10 à 14 ans. La sexualité avec un partenaire débute 2 à 4 années plus tard, entre 13 et 17 ans. Le premier coït, indicateur d'activités érotiques plus diversifiées et plus complètes, est réalisé 1 à 3 années plus tard, entre l'âge de 16 à 18 ans. Le coït anal est pratiqué plus tardivement, entre l'âge de 20 à 25 ans, et près d'un tiers des personnes ne le pratique pas. Il s'écoule en général entre 1 à 3 ans entre la date de la première réalisation d'une pratique sexuelle et la période où cette pratique devient régulière.
On observe également que les hommes et les femmes qui ont la même orientation sexuelle ont un profil similaire. De plus, les homosexuels et les bisexuels ont une sexualité plus précoce que les hétérosexuels. Les personnes les plus bisexuelles (uniquement 3, 4 et 5 sur l'échelle de Kinsey) ont la sexualité la plus précoce. Ces bisexuels ont des activités sexuelles plus tôt que les autres et le délai entre les différentes pratiques est plus court.
b – Pratiques sexuelles
Les tableaux ci-dessous indiquent la fréquence des pratiques érotiques des participants à l'enquête, en fonction du genre et de l'orientation des activités sexuelles. Les pratiques analysées sont l'échangisme, le triolisme, la sexualité de groupe et un ensemble regroupant toutes les autres pratiques supposées moins fréquentes (sado-masochisme, ondinisme, bondage, etc.).
( réponses à la question n° 9 )
|
|
Groupe (genre + orientation) |
||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
|||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
||||
|
Activité échangisme |
Non |
83 |
96,5% |
77 |
98,7% |
23 |
67,6% |
55,6% |
33 |
76,7% |
72,2% |
2 |
50,0% |
| Au - 1 x |
1 |
1,2% |
|
|
2 |
5,9% |
11,1% |
6 |
14,0% |
16,7% |
1 |
25,0% |
|
| + fois |
1 |
1,2% |
1 |
1,3% |
7 |
20,6% |
27,8% |
4 |
9,3% |
11,1% |
|
|
|
| Régulier |
1 |
1,2% |
|
|
2 |
5,9% |
5,6% |
|
|
|
1 |
25,0% |
|
|
Total |
86 |
100,0% |
78 |
100,0% |
34 |
100,0% |
N = 18 |
43 |
100,0% |
N = 18 |
4 |
100,0% |
|
Remarque : les fréquences supérieures à 5% et à 25% sont respectivement grisées claires et foncées.
|
|
Groupe (genre + orientation) |
||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
|||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
||||
|
Activité triolisme |
Non |
74 |
86,0% |
70 |
88,6% |
14 |
41,2% |
38,9% |
22 |
51,2% |
33,3% |
1 |
25,0% |
| Au - 1 x |
6 |
7,0% |
7 |
8,9% |
9 |
26,5% |
11,1% |
10 |
23,3% |
38,9% |
1 |
25,0% |
|
| + fois |
5 |
5,8% |
1 |
1,3% |
8 |
23,5% |
33,3% |
10 |
23,3% |
22,2% |
1 |
25,0% |
|
| Régulier |
1 |
1,2% |
1 |
1,3% |
3 |
8,8% |
16,7% |
1 |
2,3% |
5,6% |
1 |
25,0% |
|
|
Total |
86 |
100,0% |
79 |
100,0% |
34 |
100,0% |
N = 18 |
43 |
100,0% |
N = 18 |
4 |
100,0% |
|
Remarque : les fréquences supérieures à 15% et à 30% sont respectivement grisées claires et foncées.
|
|
Groupe (genre + orientation) |
|||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
||||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
|
||||
|
Activité sexuelle en groupe |
Non |
81 |
93,1% |
78 |
98,7% |
23 |
67,6% |
55,6% |
28 |
65,1% |
50,0% |
1 |
25,0% |
|
| Au - 1 x |
3 |
3,4% |
|
|
7 |
20,6% |
27,8% |
9 |
20,9% |
27,8% |
1 |
25,0% |
|
|
| + fois |
2 |
2,3% |
|
|
2 |
5,9% |
11,1% |
6 |
14,0% |
22,2% |
1 |
25,0% |
|
|
| Régulier |
1 |
1,1% |
1 |
1,3% |
2 |
5,9% |
5,6% |
|
|
|
1 |
25,0% |
|
|
|
Total |
87 |
100,0% |
79 |
100,0% |
34 |
100,0% |
N = 18 |
43 |
100,0% |
N = 18 |
4 |
100,0% |
|
|
Remarque : les fréquences supérieures à 10% et à 25% sont respectivement grisées claires et foncées.
|
|
Groupe (genre + orientation) |
|||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
||||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
|
||||
|
Activité sexuelle autres |
Non |
77 |
89,5% |
67 |
84,8% |
20 |
58,8% |
55,6% |
29 |
70,7% |
68,8% |
4 |
100,0% |
|
| Au - 1 x |
2 |
2,3% |
8 |
10,1% |
8 |
23,5% |
27,8% |
4 |
9,8% |
12,5% |
|
|
|
|
| + fois |
4 |
4,7% |
2 |
2,5% |
2 |
5,9% |
|
6 |
14,6% |
12,5% |
|
|
|
|
| Régulier |
3 |
3,5% |
2 |
2,5% |
4 |
11,8% |
16,7% |
2 |
4,9% |
6,3% |
|
|
|
|
|
Total |
86 |
100,0% |
79 |
100,0% |
34 |
100,0% |
N = 18 |
41 |
100,0% |
N = 18 |
4 |
100,0% |
|
|
Remarque : les fréquences supérieures à 10% et à 20% sont respectivement grisées claires et foncées.
La principale observation est que la grande majorité des personnes ne pratiquent pas régulièrement ces types d'activités érotiques. Les activités échangistes et la sexualité de groupe sont les pratiques les moins usitées, le triolisme est une pratique un peu plus fréquente, et environ 20% de l'échantillon a déjà essayé un autre type de pratique érotique. Les informations disponibles pour les homosexuels sont trop limitées pour être analysées. On observe surtout que ce sont les bisexuels, et plus les femmes que les hommes, qui pratiquent le plus les activités érotiques culturellement inhabituelles. Enfin, les personnes les plus bisexuelles (uniquement 3, 4 et 5 sur l'échelle de Kinsey) ont les activités sexuelles les plus diversifiées.
g – Nombre de partenaires
Les tableaux et figures ci-dessous indiquent le nombre total de partenaires érotiques que les sujets sélectionnés ont eu au cours de leur vie, en fonction du genre et de l'orientation des activités sexuelles.
( réponses à la question n° 7 )
|
|
Groupe (genre + orientation) |
||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
|||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
||||
|
Nombre partenaire |
1 part |
20 |
17,5% |
23 |
16,9% |
1 |
1,7% |
|
2 |
3,1% |
3,3% |
3 |
17,6% |
| 2 part |
10 |
8,8% |
18 |
13,2% |
3 |
5,2% |
6,5% |
1 |
1,6% |
3,3% |
2 |
11,8% |
|
| 3 à 5 part |
32 |
28,1% |
48 |
35,3% |
15 |
25,9% |
19,4% |
13 |
20,3% |
20,0% |
3 |
17,6% |
|
| 6 à 10 p. |
26 |
22,8% |
13 |
9,6% |
11 |
19,0% |
22,6% |
11 |
17,2% |
20,0% |
4 |
23,5% |
|
| 11 à 20 p. |
14 |
12,3% |
21 |
15,4% |
8 |
13,8% |
16,1% |
14 |
21,9% |
20,0% |
3 |
17,6% |
|
| 21 à 50 p. |
10 |
8,8% |
11 |
8,1% |
13 |
22,4% |
16,1% |
20 |
31,3% |
33,3% |
2 |
11,8% |
|
| + de 50 p. |
2 |
1,8% |
2 |
1,5% |
7 |
12,1% |
19,4% |
3 |
4,7% |
|
|
|
|
|
Total |
114 |
100,0% |
136 |
100,0% |
58 |
100,0% |
N = 31 |
64 |
100,0% |
N = 30 |
17 |
100,0% |
|
Remarque : les fréquences supérieures à 15% et à 30% sont respectivement grisées claires et foncées.
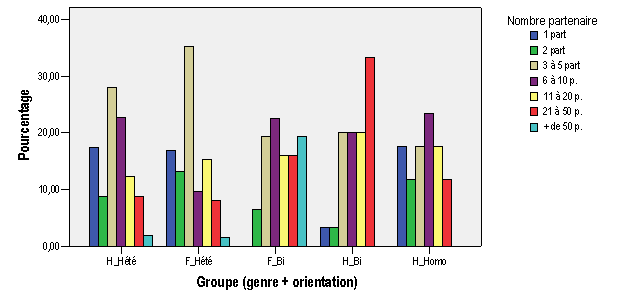
On observe que les hommes et les femmes qui ont la même orientation sexuelle ont un profil de réponse relativement similaire. La grande majorité des hétérosexuels ont entre 1 à 10 partenaires. Par contre, les homosexuels et surtout les personnes ayant des activités érotiques bisexuelles ont beaucoup plus de partenaires que les hétérosexuels.
Par ailleurs, la quasi totalité des sujets hétérosexuels de l'échantillon déclarent avoir actuellement un partenaire et en être amoureux (réponses à la question n° 5, voir tableau ci-dessous). Les bisexuels, et en particulier les femmes, sont plus nombreuses à ne pas avoir de partenaire actuellement (22,7%) et à ne pas être amoureuse quand elles en ont un (33,3%).
|
|
Groupe (genre + orientation) |
||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
|||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
||||
|
Partenaire actuellement |
Non |
3 |
4,8% |
3 |
6,4% |
5 |
22,7% |
27,3% |
5 |
13,9% |
13,3% |
|
|
| Oui |
59 |
95,2% |
44 |
93,6% |
17 |
77,3% |
72,7% |
31 |
86,1% |
86,7% |
1 |
100,0% |
|
|
Total |
62 |
100,0% |
47 |
100,0% |
22 |
100,0% |
100,0% |
36 |
100,0% |
100,0% |
1 |
100,0% |
|
|
Etat amoureux |
Non |
6 |
10,0% |
4 |
9,3% |
6 |
33,3% |
37,5% |
3 |
10,0% |
7,7% |
1 |
100,0% |
| Oui |
54 |
90,0% |
39 |
90,7% |
12 |
66,7% |
62,5% |
27 |
90,0% |
92,3% |
|
|
|
|
Total |
60 |
100,0% |
43 |
100,0% |
18 |
100,0% |
N = 8 |
30 |
100,0% |
N = 13 |
1 |
100,0% |
|
Remarque : pour les réponses négatives, les fréquences supérieures à 15% sont grisées.
d – Fréquence des activités érotiques avec un partenaire
Les tableaux et figures ci-dessous indiquent, pour chaque groupe constitué, la fréquence des activités érotiques des participants à l'enquête quand ils ont un partenaire.
( réponses à la question n° 6 )
|
|
Groupe (genre + orientation) |
||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
|||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
||||
|
Fréquence sexe avec partenaire |
- 1 x / mois |
2 |
1,8% |
4 |
2,9% |
2 |
3,4% |
6,5% |
3 |
4,8% |
6,7% |
5 |
29,4% |
| 1 x / mois |
4 |
3,5% |
|
|
2 |
3,4% |
3,2% |
4 |
6,3% |
10,0% |
2 |
11,8% |
|
| + x / mois |
14 |
12,3% |
12 |
8,8% |
3 |
5,2% |
3,2% |
9 |
14,3% |
13,3% |
|
|
|
| 1 x / sem |
10 |
8,8% |
11 |
8,1% |
5 |
8,6% |
3,2% |
5 |
7,9% |
|
4 |
23,5% |
|
| + x / sem |
52 |
45,6% |
76 |
55,9% |
24 |
41,4% |
35,5% |
27 |
42,9% |
43,3% |
3 |
17,6% |
|
| 1 x / jour |
20 |
17,5% |
15 |
11,0% |
12 |
20,7% |
25,8% |
7 |
11,1% |
6,7% |
3 |
17,6% |
|
| + x / jour |
12 |
10,5% |
18 |
13,2% |
10 |
17,2% |
22,6% |
8 |
12,7% |
20,0% |
|
|
|
|
Total |
114 |
100,0% |
136 |
100,0% |
58 |
100,0% |
N = 31 |
63 |
100,0% |
N = 30 |
17 |
100,0% |
|
Remarque : les fréquences supérieures à 15% et à 30% sont respectivement grisées claires et foncées.
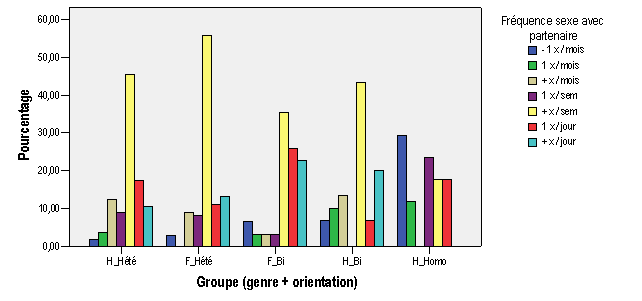
On observe que la fréquence moyenne des activités sexuelles est globalement de plusieurs fois par semaine pour tous les groupes, sauf les homosexuels qui ont une fréquence plus faible. Les hétérosexuels et les hommes bisexuels ont un profil de réponse relativement similaire. Par contre, les femmes bisexuelles, et surtout les plus bisexuelles, ont plus d'activités érotiques avec leurs partenaires que les autres groupes.
e – Fréquence des activités auto-érotiques
Les tableaux et figures ci-dessous indiquent, pour chaque groupe constitué, la fréquence des activités auto-érotique des sujets sélectionnés quand ils n'ont pas de partenaires.
( réponses à la question n° 3 )
|
|
Groupe (genre + orientation) |
||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
|||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
||||
|
Fréquence sexe mastur-bation |
- 5 fois |
2 |
1,8% |
|
|
|
|
|
|
|
|
|
|
| - 1 x / mois |
6 |
5,3% |
15 |
11,1% |
6 |
10,5% |
6,7% |
|
|
|
|
|
|
| 1 x / mois |
1 |
,9% |
8 |
5,9% |
1 |
1,8% |
3,3% |
|
|
|
|
|
|
| + x / mois |
9 |
7,9% |
32 |
23,7% |
10 |
17,5% |
20,0% |
|
|
|
|
|
|
| 1 x / sem |
22 |
19,3% |
19 |
14,1% |
11 |
19,3% |
20,0% |
4 |
6,3% |
3,3% |
1 |
5,9% |
|
| + x / sem |
43 |
37,7% |
45 |
33,3% |
19 |
33,3% |
26,7% |
28 |
43,8% |
26,7% |
9 |
52,9% |
|
| 1 x / jour |
20 |
17,5% |
13 |
9,6% |
5 |
8,8% |
6,7% |
18 |
28,1% |
36,7% |
3 |
17,6% |
|
| + x / jour |
11 |
9,6% |
3 |
2,2% |
5 |
8,8% |
16,7% |
14 |
21,9% |
33,3% |
4 |
23,5% |
|
|
Total |
114 |
100,0% |
135 |
100,0% |
57 |
100,0% |
N = 30 |
64 |
100,0% |
N = 30 |
17 |
100,0% |
|
Remarque : les fréquences supérieures à 15% et à 30% sont respectivement grisées claires et foncées.
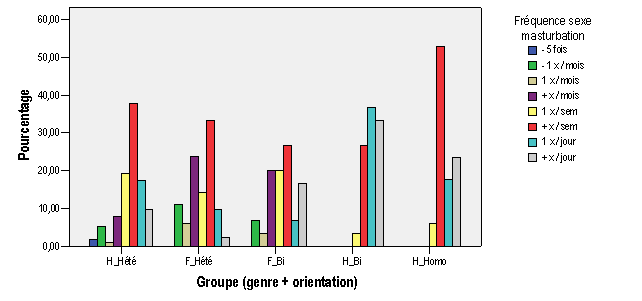
On observe que les femmes, indépendamment de l'orientation de leurs activités érotiques, se masturbent moins (environ une fois par semaine en moyenne) que les hommes. Chez les hommes, les hétérosexuels (plusieurs fois par semaine) se masturbent moins que les bisexuels et les homosexuels (près d'une fois par jour). Ce sont les personnes les plus bisexuelles, et en particulier les hommes, qui ont la plus grande fréquence d'activités auto-érotiques.
z – Période de plaisir maximal
Les tableaux et figures ci-dessous concernent, pour chaque groupe constitué, la période de la vie des participants à l'enquête où ils ont vécu les plaisirs sexuels les plus intenses.
( réponses à la question n° 14 )
|
Groupe (genre + orientation) |
Période plaisir intense - début |
Période plaisir intense - durée |
|
|
H_Hété |
Mean |
23,74 |
90,17 |
|
N |
113 |
105 |
|
|
Std. Deviation |
7,791 |
110,428 |
|
|
F_Hété |
Mean |
22,44 |
35,69 |
|
N |
135 |
121 |
|
|
Std. Deviation |
5,875 |
40,490 |
|
|
F_Bi |
Mean |
( 20,83 ) 21,50 |
( 53,46 ) 52,04 |
|
N |
56 |
50 |
|
|
Std. Deviation |
6,975 |
75,834 |
|
|
H_Bi |
Mean |
( 23,00 ) 23,32 |
( 105,63 ) 90,93 |
|
N |
60 |
57 |
|
|
Std. Deviation |
9,295 |
135,934 |
|
|
H_Homo |
Mean |
18,12 |
35,13 |
|
N |
17 |
16 |
|
|
Std. Deviation |
4,484 |
27,933 |
|
|
Total |
Mean |
22,63 |
63,42 |
|
N |
381 |
349 |
|
|
Std. Deviation |
7,275 |
93,216 |
|
Remarque : les nombres en italiques correspondent aux bisexuels stricts ( 3, 4 et 5 sur l'échelle de Kinsey).
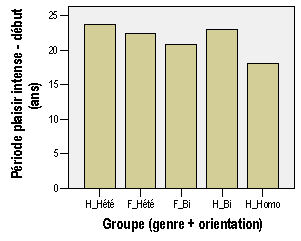
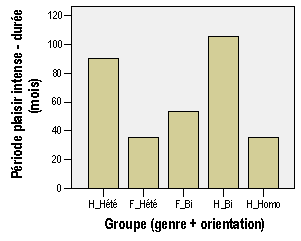
On observe que le plaisir érotique n'atteint sa pleine intensité que vers l'âge de 21-23 ans, soit 6 à 7 ans après le début des relations sexuelles. La période déclarée de plus grande intensité du plaisir érotique des hommes dure plus de deux fois plus longtemps que celle des femmes. Néanmoins cette observation est peut être un artéfact dû au fait que les hommes participants à l'enquête sont plus âgés ( 36 ans en moyenne) que les femmes (27 ans en moyenne). Les homosexuels ont leur période de plaisir intense qui débute avant les autres et les personnes les plus bisexuelles ont leur période de plaisir intense qui est plus longue que dans les autres groupes.
|
|
Groupe (genre + orientation) |
|||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
||||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
|
||||
|
Période plaisir intense-fréquence |
- 1 x / mois |
1 |
,9% |
|
|
|
|
|
1 |
1,6% |
3,6% |
1 |
5,9% |
|
| + x / mois |
5 |
4,5% |
3 |
2,2% |
3 |
5,4% |
6,7% |
1 |
1,6% |
3,6% |
1 |
5,9% |
|
|
| 1 x / sem |
7 |
6,3% |
5 |
3,7% |
1 |
1,8% |
3,3% |
2 |
3,2% |
7,1% |
4 |
23,5% |
|
|
| + x / sem |
44 |
39,3% |
68 |
50,4% |
21 |
37,5% |
30,0% |
24 |
38,7% |
21,4% |
4 |
23,5% |
|
|
| 1 x / jour |
25 |
22,3% |
23 |
17,0% |
9 |
16,1% |
16,7% |
15 |
24,2% |
25,0% |
3 |
17,6% |
|
|
| + x / jour |
30 |
26,8% |
36 |
26,7% |
22 |
39,3% |
43,3% |
19 |
30,6% |
39,3% |
4 |
23,5% |
|
|
|
Total |
112 |
100,0% |
135 |
100,0% |
56 |
100,0% |
N = 30 |
62 |
100,0% |
N = 28 |
17 |
100,0% |
|
|
Remarque : les fréquences supérieures à 20% et à 40% sont respectivement grisées claires et foncées.
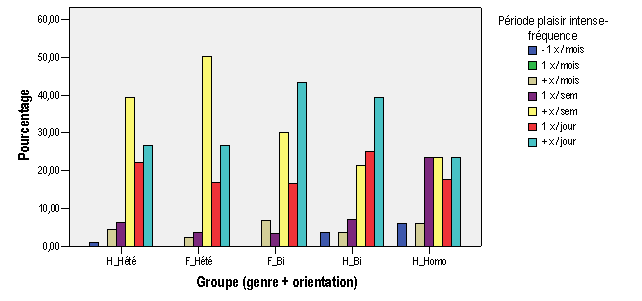
On observe que la fréquence des activités érotiques (autoérotiques et avec un partenaire) durant la période vécue de plus grand plaisir sexuel est relativement similaire entre les hommes et les femmes qui ont la même orientation sexuelle. Les homosexuels, et surtout les personnes les plus bisexuelles ont une fréquence d'activité un peu plus importante que celle des hétérosexuels.
On observe surtout que cette période où les plaisirs érotiques ont été les plus intenses est à la fois quantitative et qualitative : non seulement l'intensité du plaisir érotique de cette période est supérieur à l'intensité du plaisir des autres périodes de la vie des sujets, mais également, surtout pour les personnes les plus bisexuelles, la fréquence des activités érotiques de cette période est supérieure à la fréquence moyenne tout au long de la vie sexuelle.
h – Origine démographique
|
|
Groupe (genre + orientation) |
|||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
|
||
|
Origine démographique |
? |
2 |
1,8% |
|
|
|
|
|
|
|
|
|
| Rural |
16 |
14,0% |
12 |
8,8% |
4 |
6,9% |
11 |
17,2% |
5 |
29,4% |
|
|
| Ville < 10 |
16 |
14,0% |
31 |
22,8% |
12 |
20,7% |
10 |
15,6% |
|
|
|
|
| Ville > 10 |
80 |
70,2% |
93 |
68,4% |
42 |
72,4% |
43 |
67,2% |
12 |
70,6% |
|
|
|
Total |
114 |
100,0% |
136 |
100,0% |
58 |
100,0% |
64 |
100,0% |
17 |
100,0% |
|
|
Remarque : les fréquences supérieures à 20% sont grisées, afin de mettre en évidence le profil de l'origine démographique.
On observe que plus des 2/3 des sujets sélectionnées vivent dans des grandes agglomérations et qu'il ne semble pas exister de différence notable entre les différents groupes.
b.2 – Désir & Fantasmes
Cette section contient la description de caractéristiques plutôt psychologiques de la sexualité des sujets sélectionnés pour tester l'hypothèse HBy .
a – Désir
Les tableaux et figures ci-dessous indiquent, pour chaque groupe constitué, l'intensité du désir et la fréquence désirée des activités érotiques que les participants à l'enquête souhaiteraient avoir avec leur partenaire.
( réponses à la question n° 21 )
|
Groupe (genre + orientation) |
Mean |
N |
Std. Deviation |
|
H_Hété |
8,00 |
87 |
1,479 |
|
F_Hété |
7,92 |
77 |
1,048 |
|
F_Bi |
( 8,28 ) 8,24 |
34 |
,855 |
|
H_Bi |
( 8,29 ) 8,19 |
42 |
,994 |
|
H_Homo |
8,25 |
4 |
,957 |
|
Total |
8,05 |
244 |
1,187 |
( réponses à la question n° 11 )
|
|
Groupe (genre + orientation) |
||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
|||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
||||
|
Fréquence sexe si libre |
0 |
1 |
1,0% |
3 |
2,8% |
|
|
|
|
|
|
|
|
| - 1 x / mois |
1 |
1,0% |
1 |
,9% |
1 |
2,6% |
4,8% |
|
|
|
|
|
|
| 1 x / mois |
|
|
4 |
3,7% |
1 |
2,6% |
|
1 |
1,9% |
4,0% |
|
|
|
| + x / mois |
4 |
4,0% |
6 |
5,6% |
2 |
5,1% |
9,5% |
1 |
1,9% |
|
|
|
|
| 1 x / sem |
1 |
1,0% |
2 |
1,9% |
1 |
2,6% |
4,8% |
|
|
|
|
|
|
| + x / sem |
34 |
33,7% |
38 |
35,5% |
13 |
33,3% |
28,6% |
15 |
28,3% |
24,0% |
2 |
25,0% |
|
| 1 x / jour |
30 |
29,7% |
18 |
16,8% |
6 |
15,4% |
9,5% |
12 |
22,6% |
16,0% |
4 |
50,0% |
|
| + x / jour |
30 |
29,7% |
35 |
32,7% |
15 |
38,5% |
42,9% |
24 |
45,3% |
56,0% |
2 |
25,0% |
|
|
Total |
101 |
100,0% |
107 |
100,0% |
39 |
100,0% |
N = 21 |
53 |
100,0% |
N = 25 |
8 |
100,0% |
|
Remarque : les fréquences supérieures à 20% et à 40% sont respectivement grisées claires et foncées.
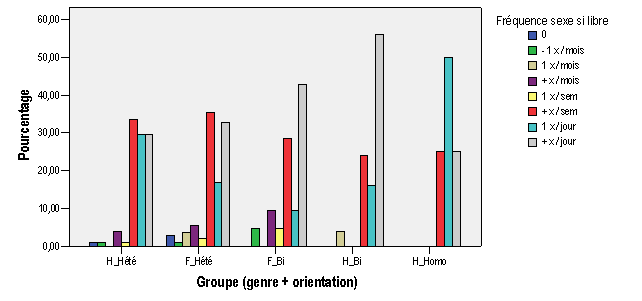
On observe que les femmes et les hommes d'un même groupe d'orientation sexuel ont un profil de réponse relativement similaire. Les informations disponibles pour les homosexuels sont trop limitées pour être analysées. Les personnes ayant des activités érotiques bisexuelles, et davantage les personnes les plus bisexuelles, ont un désir d'une plus grande fréquence d'activités érotiques avec leurs partenaires que les hétérosexuels. Les hétérosexuels souhaitent avoir des activités érotiques d'une fréquence comprise entre plusieurs fois par semaine (»35%) à plusieurs fois par jour (»30%), tandis que près de la moitié des hommes bisexuels (»50%) souhaitent avoir des relations sexuelles plusieurs fois par jour.
En comparant ces fréquences désirées avec les fréquence réelles (cf. tableau p. 187 ), on observe que tant les hétérosexuels que les bisexuels désirent augmenter leurs activités érotiques dans une proportion similaire. Même dans une situation théoriquement "libre" de toutes contraintes, les bisexuels, et surtout les personnes les plus bisexuelles, ont une sexualité plus importante que les hétérosexuels.
b – Fantasmes
Les tableaux ci-dessous indiquent, pour chaque groupe constitué, différentes caractéristiques des fantasmes érotiques des participants à l'enquête.
( réponses à la question n° 20 )
|
|
Groupe (genre + orientation) |
|||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
||||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
|
||||
|
Fréquence des fantasmes |
Rarement |
11 |
12,6% |
16 |
20,3% |
3 |
9,1% |
5,6% |
1 |
2,3% |
|
|
|
|
| Souvent |
43 |
49,4% |
48 |
60,8% |
13 |
39,4% |
27,8% |
14 |
32,6% |
33,3% |
3 |
75,0% |
|
|
| Très souv. |
33 |
37,9% |
15 |
19,0% |
17 |
51,5% |
66,7% |
28 |
65,1% |
66,7% |
1 |
25,0% |
|
|
|
Total |
87 |
100,0% |
79 |
100,0% |
33 |
100,0% |
N = 18 |
43 |
100,0% |
N = 18 |
4 |
100,0% |
|
|
Remarque : les fréquences supérieures à 20% et à 45% sont respectivement grisées claires et foncées.
|
|
Groupe (genre + orientation) |
|||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
||||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
|
||||
|
Manifestation des fantasmes |
Involontaire |
35 |
40,2% |
22 |
27,8% |
11 |
33,3% |
38,9% |
14 |
32,6% |
16,7% |
1 |
25,0% |
|
| Volontaire |
22 |
25,3% |
19 |
24,1% |
7 |
21,2% |
16,7% |
7 |
16,3% |
16,7% |
2 |
50,0% |
|
|
| Invol. & Vol. |
30 |
34,5% |
38 |
48,1% |
15 |
45,5% |
44,4% |
22 |
51,2% |
66,7% |
1 |
25,0% |
|
|
|
Total |
87 |
100,0% |
79 |
100,0% |
33 |
100,0% |
N = 18 |
43 |
100,0% |
N = 18 |
4 |
100,0% |
|
|
Remarque : les fréquences supérieures à 30% et à 50% sont respectivement grisées claires et foncées.
|
|
Groupe (genre + orientation) |
|||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
||||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
|
||||
|
Fantasme stimulant |
Partenaire |
35 |
40,7% |
30 |
38,0% |
8 |
24,2% |
16,7% |
5 |
11,6% |
11,1% |
1 |
25,0% |
|
| Autre part. |
20 |
23,3% |
19 |
24,1% |
5 |
15,2% |
16,7% |
9 |
20,9% |
22,2% |
2 |
50,0% |
|
|
| Romantique |
3 |
3,5% |
3 |
3,8% |
1 |
3,0% |
5,6% |
1 |
2,3% |
|
|
|
|
|
| Groupe |
19 |
22,1% |
17 |
21,5% |
13 |
39,4% |
50,0% |
18 |
41,9% |
44,4% |
1 |
25,0% |
|
|
| Autre |
9 |
10,5% |
10 |
12,7% |
6 |
18,2% |
11,1% |
10 |
23,3% |
22,2% |
|
|
|
|
|
Total |
86 |
100,0% |
79 |
100,0% |
33 |
100,0% |
N = 18 |
43 |
100,0% |
N = 18 |
4 |
100,0% |
|
|
Remarque : les fréquences supérieures à 20% et à 35% sont respectivement grisées claires et foncées.
|
|
Groupe (genre + orientation) |
|||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
||||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
|
||||
|
Orientation des fantasmes |
Hétéro |
76 |
90,5% |
50 |
68,5% |
8 |
24,2% |
5,6% |
14 |
32,6% |
16,7% |
|
|
|
| Bisexuel |
8 |
9,5% |
20 |
27,4% |
22 |
66,7% |
77,8% |
23 |
53,5% |
77,8% |
|
|
|
|
| Homo |
|
|
3 |
4,1% |
3 |
9,1% |
16,7% |
6 |
14,0% |
5,6% |
4 |
100,0% |
|
|
|
Total |
84 |
100,0% |
73 |
100,0% |
33 |
100,0% |
N = 18 |
43 |
100,0% |
N = 18 |
4 |
100,0% |
|
|
Remarque : les fréquences supérieures à 20% et à 40% sont respectivement grisées claires et foncées.
On observe que les bisexuels, et davantage les personnes les plus bisexuelles, ont très souvent des fantasmes liés à une sexualité intense et exubérante. Par exemple, la majorité (66,7%) des bisexuels stricts ont très souvent des fantasmes sexuels tandis que la majorité (50-60%) des hétérosexuels n'en ont que souvent ; la majorité (44-50%) des personnes les plus bisexuelles ont des fantasmes de sexualité de groupe tandis la majorité (38-40%) des hétérosexuels ont des fantasmes avec un seul partenaire. Les informations disponibles pour les homosexuels sont trop limitées pour être analysées.
On observe également que les fantasmes ont très majoritairement (entre 68 à 90%) la même orientation que les activités réelles : les hétérosexuels ont des fantasmes hétérosexuels, les bisexuels des fantasmes bisexuels et les quelques homosexuels des fantasmes homosexuels.
g – Importance de la sexualité
Les tableaux ci-dessous indiquent, pour chaque groupe constitué, l'importance accordée au plaisir érotique, à l'orgasme et à la sexualité en général.
Importance du plaisir érotique
( réponses à la question n° 15 )
|
|
Groupe (genre + orientation) |
||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
|||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
||||
|
Importance plaisir sexuel |
Peu imp. |
|
|
1 |
,9% |
|
|
|
2 |
3,7% |
4,0% |
|
|
| Moyen imp. |
3 |
3,0% |
5 |
4,7% |
2 |
5,0% |
|
3 |
5,6% |
4,0% |
|
|
|
| Important |
34 |
33,7% |
39 |
36,4% |
11 |
27,5% |
19,0% |
7 |
13,0% |
16,0% |
6 |
66,7% |
|
| Très imp. |
64 |
63,4% |
62 |
57,9% |
27 |
67,5% |
81,0% |
42 |
77,8% |
76,0% |
3 |
33,3% |
|
|
Total |
101 |
100,0% |
107 |
100,0% |
40 |
100,0% |
N = 21 |
54 |
100,0% |
N = 25 |
9 |
100,0% |
|
Remarque : les fréquences supérieures à 30% et à 70% sont respectivement grisées claires et foncées.
Importance de l'orgasme
( réponses à la question n° 15 )
|
|
Groupe (genre + orientation) |
|||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
||||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
|
||||
|
Importance plaisir orgastique |
Sans imp. |
1 |
,9% |
1 |
,7% |
1 |
1,7% |
3,2% |
|
|
|
|
|
|
| Peu imp. |
3 |
2,6% |
3 |
2,2% |
|
|
|
1 |
1,6% |
|
1 |
5,9% |
|
|
| Moyen imp. |
30 |
26,3% |
35 |
25,7% |
11 |
19,0% |
12,9% |
11 |
17,2% |
23,3% |
5 |
29,4% |
|
|
| Important |
49 |
43,0% |
69 |
50,7% |
27 |
46,6% |
48,4% |
24 |
37,5% |
30,0% |
9 |
52,9% |
|
|
| Très imp. |
31 |
27,2% |
28 |
20,6% |
19 |
32,8% |
35,5% |
28 |
43,8% |
46,7% |
2 |
11,8% |
|
|
|
Total |
114 |
100,0% |
136 |
100,0% |
58 |
100,0% |
N = 31 |
64 |
100,0% |
N = 30 |
17 |
100,0% |
|
|
Remarque : les fréquences supérieures à 25% et à 40% sont respectivement grisées claires et foncées.
Importance de la sexualité
( réponses à la question n° 22 )
|
|
Groupe (genre + orientation) |
|||||||||||||
| H_Hété |
F_Hété |
F_Bi |
H_Bi |
H_Homo |
||||||||||
| N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
N |
Col % |
|
||||
|
Importance de la sexualité |
Imp. second |
4 |
3,5% |
5 |
3,7% |
|
|
|
1 |
1,6% |
|
1 |
5,9% |
|
| Imp. égale |
10 |
8,8% |
13 |
9,6% |
9 |
15,5% |
16,1% |
4 |
6,3% |
6,7% |
1 |
5,9% |
|
|
| Relativ. imp. |
20 |
17,5% |
34 |
25,0% |
10 |
17,2% |
16,1% |
7 |
10,9% |
16,7% |
7 |
41,2% |
|
|
| Grande imp. |
64 |
56,1% |
74 |
54,4% |
31 |
53,4% |
48,4% |
41 |
64,1% |
66,7% |
7 |
41,2% |
|
|
| Imp.centrale |
16 |
14,0% |
10 |
7,4% |
8 |
13,8% |
19,4% |
11 |
17,2% |
10,0% |
1 |
5,9% |
|
|
|
Total |
114 |
100,0% |
136 |
100,0% |
58 |
100,0% |
N = 31 |
64 |
100,0% |
N = 30 |
17 |
100,0% |
|
|
Remarque : les fréquences supérieures à 25% et à 60% sont respectivement grisées claires et foncées.
Conclusion
On observe que le plaisir érotique est important pour environ 95% des participants à l'enquête, très important pour les 2/3 d'entre eux, et très important pour les 3/4 des personnes les plus bisexuelles. Paradoxalement, l'orgasme, qui est pourtant l'état paroxystique du plaisir érotique, est jugé moins important par tous les groupes. Est-ce une rationalisation du fait que l'orgasme est un état très court et incontrôlable ? Quant à la sexualité en général, elle a une grande importance pour la moitié des sujets. Les réponses à ces trois questions sont similaires pour les hommes et les femmes hétérosexuels. Les bisexuels accordent une plus grande importance à ces différents aspects de la sexualité. Les hommes bisexuels se détachent assez nettement des quatre autres groupes, en donnant aux différents aspects de la sexualité une très grande importance dans leur existence.
b.3 – Conclusion
Pour les sujets sélectionnés, le schéma général de leur sexualité est le suivant. L'entrée dans la sexualité a été progressive, s'étendant entre les âges de 10 à 20 ans. Les bisexuels et les hommes ont une sexualité plus précoce et des activités plus fréquentes : les hommes bisexuels ont en moyenne environ une dizaine de partenaires avec lesquels ils ont des activités érotiques une fois par jour ; les femmes hétérosexuelles ont en moyenne environ cinq partenaire avec lesquels elles ont des rapports sexuels plusieurs fois par semaine. Les femmes débutent leur sexualité plus tard, mais ont des partenaires plus tôt que les hommes.
La grande majorité des hétérosexuels (80%) ont surtout des activités érotiques classiques en couples. La moitié des bisexuels pratiquent des activités érotiques inhabituelles, telle la sexualité de groupe. Les fantasmes correspondent globalement aux activités érotiques pratiquées.
Les homosexuels constituent ont également un profil distinct des deux autres groupes. Néanmoins, la taille limitée de l'échantillon ne permet pas des analyses fiables.
Bien que cela ne soit pas l'objet de cette étude, il semble que la différence de profil provienne surtout du fait que les homosexuels et les bisexuels font partie de groupes sociologiques qui intègre moins les normes sociales dominantes. Vraisemblablement, il ne s'agirait pas d'une différence physiologique, mais culturelle.
En raison de ces profils distincts, les différents groupes ne sont pas regroupés en un seul pour les analyses statistiques et la vérification des hypothèses.
( Cf. également la section "Participants" du chapitre "3 – Matériel & Méthode", à la page 65 )
2 – Annexe du chapitre "Résultats"
a – Le plaisir érotique est-il un des plaisirs les plus intenses que l'organisme peut éprouver ?
Dans les deux premières versions du questionnaire, la consigne pour la question n° 1 était d'indiquer les activités préférées, par ordre de préférence, avec les chiffres 1, 2, 3, etc. L'évaluation par les sujets (104 femmes et 71 hommes) des différentes activités qu'ils préfèrent a donné les résultats suivants :
Groupe : HOMMES hétérosexuels : Activités préférées
[ N = 37 ] Rang : 1er 2e 3e 4e 5e (absence de notes)
Activité sexuelle avec partenaire 33 [17] 4 (0)
Activité gustative 1 7 10 1 (18)
Activité sexuelle avec soi-même 8 7 1 (21)
Activité corporelle 6 6 1 (23)
Activité intellectuelle 1 3 4 1 1 (26)
Activité auditive 4 4 (28)
Activité visuelle 1 1 2 1 (31)
Activité olfactive 1 2 (33)
Activité artificielle 1 (35)
Groupe : Hommes bisexuels : Activités préférées
[ N = 21 ] Rang : 1er 2e 3e 4e 5e (absence de notes)
Activité sexuelle avec partenaire 17 3 1 (0)
Activité auditive 1 4 4 (12)
Activité corporelle 2 2 2 (15)
Activité gustative 3 5 (13)
Activité sexuelle avec soi-même 4 2 (15)
Activité visuelle 1 1 1 (18)
Activité intellectuelle 2 2 (17)
Activité olfactive 1 2 (18)
Activité artificielle 1 1 (19)
Groupe : HOMMES homosexuels : Activités préférées
[ N = 13 ] Rang : 1er 2e 3e 4e 5e (absence de notes)
Activité sexuelle avec partenaire 6 3 2 (2)
Activité auditive 2 6 3 (2)
Activité intellectuelle 3 1 (9)
Activité sexuelle avec soi-même 2 1 (10)
Activité visuelle 1 2 (10)
Activité artificielle 2 (11)
Activité gustative 1 (12)
Activité corporelle 1 (12)
Activité olfactive 1 (12)
Groupe : FEMMES hétérosexuelles : Activités préférées
[ N = 80 ] Rang : 1er 2e 3e 4e 5e (absence de notes)
Activité sexuelle avec partenaire 67 6 2 (5)
Activité gustative 8 15 18 2 (35)
Activité corporelle 19 9 1 3 (47)
Activité auditive 1 14 14 1 1 (46)
Activité intellectuelle 3 5 13 1 (54)
Activité sexuelle avec soi-même 1 10 7 (58)
Activité visuelle 5 6 3 (62)
Activité olfactive 1 5 1 2 (68)
Activité artificielle 2 2 1 (73)
Groupe : Femmes bisexuelles : Activités préférées
[ N = 24 ] Rang : 1er 2e 3e 4e 5e (absence de notes)
Activité sexuelle avec partenaire 16 3 2 (3)
Activité corporelle 3 1 5 (15)
Activité intellectuelle 2 4 1 (17)
Activité gustative 5 4 (15)
Activité sexuelle avec soi-même 1 4 2 (17)
Activité auditive 1 3 4 (16)
Activité olfactive 2 2 (20)
Activité visuelle 1 1 (22)
Activité artificielle 1 (23)
Conclusion
Tant chez les hommes que chez les femmes, quel que soit l'orientation des activités sexuelles, on observe que l'activité érotique avec un partenaire est, de très loin, l'activité préférée.
La principale différence entre les deux protocoles de notation est l'existence ou non d'ex aequo dans l'ordre de préférence des activités. Les deux premières versions du questionnaire ont accentué la hiérarchie entre les préférences, tandis que la troisième a peut être atténué cette différence.
Mais quel que soit le système de notation, on retrouve le même résultat, qui est la nette préférence pour les activités érotiques avec un partenaire.
b – Les zones érogènes des organes génitaux sont-elles à l'origine du plaisir érotique le plus intense ?
Dans les deux premières versions du questionnaire, la consigne pour la question n° 17 était d'indiquer les activités sexuelles préférées, par ordre de préférence, avec les chiffres 1, 2, 3, etc. L'évaluation par les sujets (104 femmes et 71 hommes) des différentes activités érotiques qu'ils préfèrent donne les résultats suivants :
Groupe : Hommes hétérosexuels : Activités sexuelles préférées
[ N = 37 ] Rang : 1er 2e 3e 4e 5e (absence de notes)
Coït vaginal 16 7 6 (8)
Fellation passif 5 9 3 2 (17)
Cunnilingus 7 4 4 1 1 (20)
69 1 5 7 1 (23)
Coït anal 3 3 1 (29)
Embrasser 2 2 2 (29)
Masturbation réciproque 1 6 (29)
Masturbation passif 1 2 (32)
Contact corporel 2 1 (31)
Masturbation actif 1 (35)
Caresses sensuelles 1 (35)
Regroupement des activités sexuelles par zones érogènes stimulées :
Pénis stimulé ( Coït vaginal + Coït anal + Fellation passif + Masturbation passif )
25 19 12 2
Oral stimulé ( Embrasser + Cunnilingus )
9 4 6 1 3
Cunnilingus 7 4 4 1 1
69 1 5 7 1
Contact corporel 2 1
On observe que chez les hommes hétérosexuels le pénis est, de loin, la zone érogène autour de laquelle se réalisent les activités érotiques.
Groupe : Femmes hétérosexuelles : Activités sexuelles préférées
[ N = 80 ] Rang : 1er 2e 3e 4e 5e (absence de notes)
Coït vaginal passif 27 18 13 1 2 (18)
Cunnilingus passif 18 12 8 1 (36)
Fellation actif 7 12 8 3 (46)
Contact corporel 12 5 6 2 1 (52)
69 2 10 7 (56)
Masturber passif 4 7 6 1 1 (57)
Embrasser 5 3 10 2 (56)
Masturbation réciproque 1 5 7 1 (61)
Caresses sensuelles 1 2 3 2 (69)
Coït anal passif 2 5 1 (65)
Fesse passif 4 1 (72)
Fesse actif 1 1 (74)
Masturbation actif 1 1 (72)
Regroupement des activités sexuelles par zones érogènes stimulées :
Clitoris + Coït vaginal passif 50 42 34 3 4
Clitoris (total) ( cunnilingus passif + masturbation (passif + réciproque) )
23 24 21 2 2
Coït vaginal passif 27 18 13 1 2
Oral (total) ( embrasser + fellation actif )
12 15 18 3 2
Contact corporel 12 5 6 2 1
69 2 7 6
Région anale ( Coït anal passif + Fesse passif )
2 9 2
Chez les femmes hétérosexuelles, le clitoris, le vagin et la zone orale sont les 3 régions autour desquelles se réalisent les activités érotiques.
Dans la mesure où le clitoris est également stimulé lors du coït vaginal (de manière externe et de manière interne), on observe que le clitoris est, de loin, la zone érogène autour de laquelle se réalisent les activités érotiques.
Groupe : Femmes bisexuelles : Activités sexuelles préférées
[ N = 24 ] Rang : 1er 2e 3e 4e 5e (absence de notes)
Coït vaginal passif 9 1 5 (9)
Cunnilingus passif 3 3 2 (16)
Contact corporel 3 2 2 (17)
Fellation actif 1 6 3 (14)
Embrasser 3 2 1 (18)
Masturber passif 3 1 1 (19)
Coït anal passif 3 4 (17)
69 2 3 (19)
Anus passif 1 (23)
Fesse actif 1 (23)
Masturber réciproque 1 (23)
Caresses sensuelles 1 (23)
Cunnilingus actif 2 (22)
Regroupement des activités sexuelles par zones érogènes stimulées :
Clitoris + Coït vaginal passif 15 5 8
Coït vaginal passif 9 1 5
Oral (total) ( embrasser + fellation actif )
4 8 4
Clitoris (total) ( cunnilingus passif + masturbation passif )
6 4 3
Contact corporel 3 2 2
Chez les femmes bisexuelles, le clitoris, le vagin et la zone orale sont les 3 régions autour desquelles se réalisent les activités érotiques.
Dans la mesure où le clitoris est également stimulé lors du coït vaginal (de manière externe et de manière interne), on observe que le clitoris est, de loin, la zone érogène autour de laquelle se réalisent les activités érotiques.
Groupe : Hommes bisexuels : Activités sexuelles préférées
[ N = 21 ] Rang : 1er 2e 3e 4e 5e (absence de notes)
Coït vaginal 6 4 2 (9)
69 4 2 2 (13)
Cunnilingus 2 1 5 (13)
Fellation passif 1 4 1 (15)
Coït anal actif 1 2 2 (16)
Fellation actif 2 1 1 (17)
Contact corporel 1 2 1 (17)
Masturbation passif 1 1 2 (17)
Masturbation réciproque 1 1 1 (18)
Coït anal passif 1 1 (19)
Embrasser 1 2 (18)
Fesse passif 1 (20)
Fesse actif 1 (20)
Masturbation actif 1 (20)
Caresses sensuelles 1 (20)
Regroupement des activités sexuelles par zones érogènes stimulées :
Pénis stimulé (Total) ( [ pénis stimulé ]+ Masturber Reciproque + 69 )
14 14 10
Pénis stimulé ( Coït vaginal + Coït anal actif+ Fellation passif + Masturb. passif )
9 11 7
Oral stimulé (Total) ( [ Oral stimulé ] + 69 )
8 5 10
Oral stimulé ( Embrasser + Cunnilingus + Fellation actif )
4 3 8
Manuel actif (Total) ( [ Manuel actif ] + Masturbation réciproque )
1 2 3
Corps stimulé ( Contact corporel + Caresses sensuelles )
1 2 2
Zone anale passif ( Coït anal passif + fesse passif )
2 1
Manuel actif ( Fesse actif + Masturbation actif + Caresses sensuelles )
1 2
On observe que chez les hommes bisexuels le pénis (principalement) et la zone orale (secondairement) sont les 2 zones érogènes autour desquelles se réalisent les activités érotiques.
Groupe : Hommes homosexuels : Activités sexuelles préférées
[ N = 13 ] Rang : 1er 2e 3e 4e 5e (absence de notes)
Contact corporel 5 1 1 (6)
Fellation actif 3 2 3 (5)
Coït anal passif 2 1 2 (8)
Embrasser 1 3 1 (8)
Fellation passif 1 1 2 (11)
Masturbation réciproque 1 2 (10)
Coït anal actif 1 1 (11)
69 1 1 (11)
Masturbation passif 3 (10)
Fesse actif 3 (12)
Masturbation actif 1 (12)
Regroupement des activités sexuelles par zones érogènes stimulées :
Oral stimulé (Total) ( [ Oral stimulé ] + 69 )
4 6 5
Oral stimulé ( Embrasser + Fellation actif )
4 5 4
Pénis stimulé (Total) ( [ pénis stimulé ]+ Masturber Reciproque + 69 )
3 5 6
Corps stimulé ( Contact corporel + Caresses sensuelles )
5 1 1
Pénis stimulé ( Coït anal actif+ Fellation passif + Masturbation passif )
2 2 5
Zone anale passif ( Coït anal passif + fesse passif )
2 1 2
Manuel actif (Total) ( [ Manuel actif ] + Masturbation réciproque )
1 3
Manuel actif ( Fesse actif + Masturbation actif + Caresses sensuelles )
1 3
La principale différence entre les deux protocoles de notation est l'existence ou non d'ex aequo dans l'ordre de préférence des activités sexuelles. Les deux premières versions du questionnaire ont accentué la hiérarchie entre les préférences, tandis que la troisième a peut être amoindri cette différence.
On observe que chez les hommes le pénis est une zone érogène autour de laquelle se réalisent les activités érotiques. Mais les contacts corporels non sexuels et les activités où le sujet stimule son partenaire sont également des activités importantes, contrairement à ce que prévoit le modèle théorique.
Conclusion
On observe dans tous les groupes, sauf pour les homosexuels, que le pénis/clitoris est la principale zone érogène autour de laquelle s'organisent les activités érotiques. La zone orale est la seconde zone érogène organisatrice des activités érotiques. Le coït vaginal est l'activité préférée, mais plusieurs autres activités érotique sont également fréquemment pratiquées.
Annexe IV : Problèmes relatifs aux expériences éthiquement réalisables
Cette annexe contient la présentation d'une des différentes observations qui ont été tentées afin de tester les hypothèses de cette recherche. La plupart de ces vérifications expérimentales ont été abandonnées en raison de problèmes similaires à ceux décrits ci-dessous.
Une des possibilités de vérifier l'absence chez l'Homme d'instinct de la reproduction est de rechercher dans la population générale des couples hétérosexuels ayant eu des activités sexuelles tout en ignorant l'existence du coït vaginal. Il semble même possible de trouver une situation encore plus intéressante, lorsque, de surcroît, au moins une des deux personnes du couple savait, par déduction d'informations partielles et/ou implicites, qu'il existait une activité sexuelle particulière à effectuer au cours des activités sexuelles.
L'observation a posteriori des activités érotiques de plusieurs de ces couples, par reconstitution après une interview ou un questionnaire, devrait permettre de tester cette hypothèse de l'absence d'instinct.
En effet, dans les conditions de base – où les partenaires sont nus, se stimulent physiquement et sont dans un état d'excitation sexuelle – s'il existe des processus innés permettant la réalisation du coït vaginal, il est fortement probable que le coït soit réalisé. Dans ce cas, si on observe que la majorité des couples réalisent le coït vaginal, l'hypothèse de l'absence d'instinct peut être considérée comme réfutée.
Dans le cas où le couple ne réalise pas le coït vaginal, surtout si une ou les deux personnes savent qu'il faut réaliser un acte sexuel particulier, il est vraisemblable qu'il n'existe pas de processus innés du coït vaginal. Dans ce cas, si on observe que la majorité des couples ne réalisent pas le coït vaginal, l'hypothèse de l'absence d'instinct peut être considérée comme validée.
Cette situation "naturelle" semble être la meilleure que l'on puisse trouver – en particulier dans le cadre de cette thèse – afin de réfuter ou de valider cette hypothèse à partir de données humaines.
Étude de cas
Dans un premier temps, un réseau de sexologue a été sollicité pour trouver dans leur clientèle des personnes ayant été dans cette situation particulière. À ce jour, seule une personne ayant eu une activité hétérosexuelle tout en ignorant le coït vaginal a pu être trouvée et questionnée. Son cas est présenté ci-dessous.
AP a 52 ans. Il est originaire de Pologne, d'un milieu rural très catholique. La sexualité était un sujet tabou et à sa majorité AP n'avait que très peu d'expériences et d'informations sexuelles.
À 17 ans, il vit sa première expérience sexuelle avec une adolescente de 19 ans. Ils sont couchés dans un lit, se caressent, s'embrassent et se déshabillent. AP sait, par des bribes d'informations provenant de ses camarades, qu'il doit «faire quelque chose». Il est sexuellement excité, en érection. À un moment, l'adolescente l'enserre dans ses bras et AP se couche sur elle. Leur sexes sont en contact. Il n'y a pas de pénétration. Après quelques minutes, AP ressent un "blocage" : «je n'étais pas à l'aise», et met fin à l'activité sexuelle.
Ce cas, a priori, puisque le coït vaginal n'a pas été réalisé malgré les contacts physiques adéquats et l'état d'excitation sexuelle, valide l'hypothèse de la non-innéité du coït vaginal.
Néanmoins, il se dégage de l'entretien avec AP que les deux adolescents n'étaient pas à l'aise : quasiment aucun échange verbal, peu d'activités érotiques, pas d'éjaculation ni d'orgasme, et "blocage" de l'activité sexuelle après une dizaine de minutes. Bien qu'aucun problème particulier n'ait été évoqué (peur de ne pas être à la hauteur, peur de tomber enceinte, vaginisme, etc.), on perçoit clairement, sans pouvoir préciser la nature exacte du problème, que la situation sexuelle était loin d'être optimale.
Problèmes
En fait, il existe plusieurs problèmes qui rendent difficile l'exploitation de cette situation sociale et culturelle particulière, et qui, de ce fait, limitent la portée des observations qui pourraient être réalisées.
Dans un premier temps, il est très difficile de trouver des couples hétérosexuels ayant été dans l'ignorance du coït vaginal. D'une part, depuis la révolution culturelle des années 70, la diffusion de l'information sexuelle rend quasi improbable le fait de trouver un couple où les deux partenaires ignorent le coït vaginal. D'autre part, dans la population des plus de 50 ans où il semble possible de trouver quelques cas de ce type, il faudrait pouvoir contacter des dizaines de milliers de personnes pour trouver les rares personnes qui correspondent aux critères expérimentaux et qui accepteront de témoigner. Cela demande une logistique et des moyens non disponibles dans le cadre de cette thèse.
Ensuite, il est possible qu'il existe des apprentissages latents, non conscients, par exemple par des observations de copulation d'animaux. Généralement, ces apprentissages non explicites ne peuvent être détectés et leur existence potentielle rend ainsi inexploitables les résultats de l'observation.
De surcroît, basé sur l'expérience clinique en sexologie, il semble qu'en général si des personnes n'ont pas reçu avant la puberté d'informations sur le coït vaginal, c'est en raison d'un contexte familial et/ou culturel défavorable à la sexualité. Dans ces contextes, on observe qu'en général le problème n'est pas limité à la simple censure d'informations sexuelles, mais correspond à un état psychologique particulier par rapport à la sexualité : tout ce qui relève de la sphère sexuelle est plus ou moins tabou, et est accompagné de réactions particulières, de non-dits et de réactions émotionnelles plutôt négatives. Pour des personnes ayant vécus dans de tels contextes, l'évocation des souvenirs sexuels est souvent gênante et pénible, et des éléments sensibles peuvent être occultés ou dissimulés. De plus, les personnes peuvent présenter des problèmes plus ou moins importants et divers en rapport avec la sexualité (ignorance, honte, dégoût des parties génitales, éjaculation précoce, frigidité, ...). Tous ces problèmes permettent de supposer l'existence de divers facteurs psychologiques qui empêcheraient la libre expression des éventuels processus innés permettant le coït vaginal. Pour ces raisons, dans les cas où le coït vaginal n'est pas réalisé, il est toujours possible de supposer l'intervention par exemple d'une inhibition psychique et il est donc quasiment impossible d'affirmer avec certitude l'absence de processus innés.
Pour toutes ces raisons, il n'a pas été jugé pertinent de continuer cette enquête.
Liste des publications
Les travaux présentés dans ce travail ont donné lieu aux publications suivantes :
1) Publications à comité de lecture
I. Wunsch S, Brenot P (2004) Does a sexual instinct exist ? -- Sexologies, 13(48):30-36
II. Wunsch S, Brenot P (2004) Neurobiology of pleasure -- Sexologies, 13(50):17-27
III. Wunsch S, Brenot P (2005) Sexualité : instinct ou apprentissage ?, Médecine Sexuelle, 1:12-21
IV. Wunsch S, Brenot P (2005) Analyse des rapports entre structure biologique et sexualité, NeuroPsy News, 4(4):133-136
V. Wunsch S, Brenot P (2005) Un modèle synthétique des dysfonctions et pathologies sexuelles, NeuroPsy News, 4(5):162-165
VI. Wunsch S, Brenot P (2007) The role and importance of reinforcement processes in the learning of reproduction behavior in man., en préparation. pour Archives of Sexual Behavior
VII. Wunsch S, Brenot P (2007) Primordial factors of reproduction behavior and their evolution in man, en cours de re-soumission.
2) Communications orales
I. Wunsch S. Déterminants neurobiologiques du comportement sexuel -- Communication, Congrès de sexologie AIHUS, Marseille, 2006
[1] À noter que l'existence chez la femme d'un phénomène similaire à l'éjaculation masculine est controversée.
[2] Ces explications, qui auraient logiquement dû être présentées dans cette section, ont déjà été données lors de l'étude des différents systèmes sensoriels. Pour la clarté des explications, chaque système sensoriel, du récepteur au néocortex, a été étudié dans une seule section plutôt que dans plusieurs.
[3] Définition du schéma corporel : "édifié sur les impressions tactiles, kinesthésiques, labyrinthiques et visuelles, le schéma corporel réalise dans une construction active constamment remaniée des données actuelles et du passé, la synthèse dynamique, qui fournit à nos actes, comme à nos perceptions, le cadre spatial de référence où ils prennent leur signification." De AJURIAGUERRA 1970
[4] Stephen Jay Gould (1991) à également critiqué l'importance démesurée accordée à l'effet d'optimisation : «La théorie de l'évolution, telle qu'elle est présentée dans de nombreux textes de vulgarisation, attribue à la sélection naturelle le rôle d'un principe de perfection qui opère avec tant de précision et d'omnipotence que les animaux paraissent répondre à un ensemble de mécanismes incorporés, programmés pour leur faire atteindre une forme optimale».
[5] Pour exemple, une version inactivée du questionnaire est toujours accessible à cette adresse.
[6] Test non paramétrique de comparaison de plusieurs moyennes de rang pour des échantillons appariés.
[7] Test non paramétrique de comparaison de deux moyennes de rang pour des échantillons appariés.
[8] Test non paramétrique post hoc utilisé pour les comparaisons 2 à 2 entre plusieurs moyennes de rang.
[9] Test non paramétrique utilisé pour la comparaison de 2 moyennes de rang, provenant d'échantillons indépendants et constitués de données ordinales.
[10] Test non paramétrique de comparaison de plusieurs moyennes de rang pour des échantillons appariés.
[11] Test non paramétrique de comparaison de deux moyennes de rang pour des échantillons appariés.
[12] Test non paramétrique post hoc utilisé pour les comparaisons 2 à 2 entre plusieurs moyennes de rang.
[13] Test non paramétrique utilisé pour la comparaison de 2 moyennes de rang, provenant d'échantillons indépendants et constitués de données ordinales.
[14] Test non paramétrique de comparaison de plusieurs moyennes de rang pour des échantillons appariés.
[15] Test non paramétrique de comparaison de deux moyennes de rang pour des échantillons appariés.
[16] Voir les travaux pionniers de K. Lorenz et de N. Tinbergen (L'étude de l'instinct, paris, 1953).
[17] Sur les 37 personnes de ce groupe ayant répondu à cette question, 33 ont mis l'activité sexuelle avec un partenaire en premier rang et 4 en deuxième rang ; comme toutes les personnes ont répondu à cet item, il ne manque aucune note.