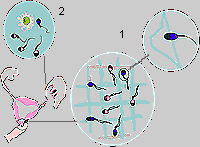|
The Theory of Pre-zygotic Sex Determination |
|||||||||||||||||||||
|
1. Introduction 1. IntroductionHuman reproductive behavior is studied by specialists of
many disciplines. Until now, however, there has been no
convincing explanation of the fall in birth-rate in developed
countries. Demographic investigations show that many modern
couples are limiting themselves to one child, even as men
and women express in opinion surveys the desire for larger
families. This discrepancy has been explained by the fact
that more than 50% of respondents said they would prefer
a family with two children, one boy and one girl. That is
to say that spouses would be more willing to have a second
child, on condition that they could be assured of having
either a son or a daughter. The impossibility of determining
the sex of a future baby has thus proved a negative influence,
decreasing the likelihood that couples will decide to have
a second child.
The possibility of regulating the gender of a child before conception has been of interest to human- kind since the dawn of time. We know of many recommendations, drawn from the sexual cultures of many nations, for how a woman might choose the sex of her baby. Of course, many of these recommendations should be met with skepticism – they are based more on mysticism than on scientific knowledge. The fact remains, however, that they are taken seriously by many people even today. Unfortunately, the current theme has not been the subject of broad scientific investigation, and any progress towards its resolution has invariably led to heated discussions, reflecting the polarity of opinions on the only 100%-effective means of controlling the birth-rate. Some people have worried that artificial regulation of a child’s gender may lead to social and demographic problems. Some authors felt that the disruption of the more-or-less stable proportion of male to female births could harmfully affect the rate of reproduction of the population (4.c. 140), or even lead to social up-heaval (5). The counter to this view is the confirmation (on the basis of sociological studies of reproductive attitudes) that an effective method of gender-determination will not lead to a disruption in the secondary sex ratio (6, c. 75). Perhaps this is really the case, because the majority of respondents express a preference for families with one boy and one girl. So, if we provide parents with the opportunity to realize their reproductive rights, there will not be any great change in the ratio of the sexes within the population. The population itself, however, may well be affected, increasing in countries where couples had previously limited themselves to one child, and decreasing in those where people usually have more than two. Few specialists doubt that the gender of a child is at least partly determined by non-random factors (in fairness, it must be noted that some have disputed this in doctoral dissertations - 7). Investigations in various branches of science have provided data on the planning of a child's gender, and methods for gender planning have been proposed (8, 9, 10). None of these theories or methods, however, gives a convincing explanation of why the process of pre-zygotic determination should exist in nature. The data show that this process has an effect on the population, and that it is a natural consequence of the division of a species into two genders. No single scientific discipline can provide sufficiently exhaustive evidence to convincingly demonstrate the biological advantage of this phenomenon. As a result, a multi-disciplinary investigation is necessary. First, we will clarify the influence of immunological factors. 2. Pre-zygotic selectionImmunological factors doubtlessly have a great effect on conception. We have evidence of this fact from the investigation of rabbits, immunized with spermatozoid antigens. The effect of the antigens drastically altered the gender-ratio among descendents along the female line. When mating took place after the introduction of X-sperm antigens, more males were born, and when Y-sperm antigens were introduced, more females. In the immunology of reproduction it is well established that significant differences exist in the characteristics of sperm carrying the different chromosomes (references 12,13,14). The protein on the surface of male sex-cells has different characteristics in X- and Y-carrying sperm. The particular characteristics of this membrane are genetically controlled, as indeed are all the other characteristics of spermatozoids. On the other hand, some factors, such as sperm's speed of motion are dependant on the specifics of the surrounding environment. On the basis of the functional and morphological differences between X- and Y-carrying sperm cells, some methods have been proposed for distinguishing between the two, with the goal of determining the sex of a baby by artificial insemination. The movement of sperm along the sexual channels before and after ejaculation are accompanied by complex immunological phenomena which are known as 'pre-zygotic selection'. Pre-zygotic selection is seen as the combination of two processes which occur in the woman's sexual tract after ejaculation. Firstly (while sperm move through the cervical filter), genetically defective sperm are eliminated by immunological reactions. After this 'rejection of the defective,' there follows 'selection of the fittest.' The essential feature of this last process in that from the pool of genetically viable sperm, only the 'best' (according to certain functional characteristics), are allowed to move towards the egg for fertilization.
Diagram 1: Pre-zygotic selection:
1 - "rejection of the defective"; 2 - "selection
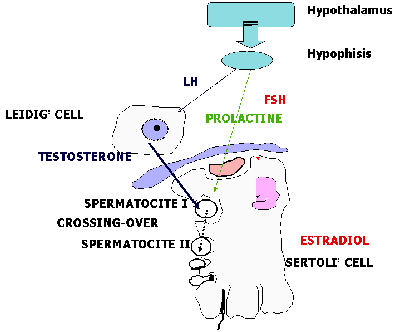
of the fittest" It is by no means impossible that the chromosome carried by a sperm-cell (i.e. X or Y), could have an effect on its selection or rejection. The theoretical positions of reproductive immunology, however, do not explain how exactly X- or Y- carrying sperm could acquire such a characteristic. To find the explanation we must turn to ideas about the regulation of spermatogenesis, and especially to the genetic phenomenon of meiotic drift. 3. Meiotic DriftIt is well established in cyto-genetics, that there is an active process which leads to genetic drift within a population. This happens during meiosis as a result of crossing-over, when during the exchange of chromosomes, factors distribute themselves in ratios different from those postulated by Mendel's laws. Experiments on fruit-flies have shown how meiotic drift can be induced (for example, by spirochaeta infection, and by certain mutagens), and to what extent it can lead to a change in the ratio of genders (in some cases, the percentage of female descendents along the male line can be as high as 95%). Crossing-over occurs at those phases of meiosis where cell-division is declining - in the stage where spermatocyte-1 transforms into spermatocyte-2 (diagram 2). It is well known that these very spermatocytes are the target cells for testosterone. Crossing-over, then, is a testosterone-dependant process. The level of testosterone production by Glandulocytes (Leidig cells), which have direct functional links to cells of the sperm-generating epithelium, is determined by genetic factors and by the intensity of sexual life.
Diagram 2: The Cytogenetic Mechanism of pre-zygotic sex determination 4. The Cytogenetic Basis of pre-zygotic sex determinationIn accordance with the above, the cyto-genetic basis of pre-zygotic gender determination can be seen as a testosterone dependant process - crossing-over during the course of meiosis. During this process, the distribution and exchange of sex-chromosomes occurs according to Mendel's laws, but the determination of the functional characteristics of X- and Y-carrying spermatozoids does not. As a result of meiotic drift, the surface membranes of X- and Y-carrying sperm acquire special characteristics which are realized during immunological reactions after the sperm enter the woman's genital tract. In the male organism, then, there is a biological 'tool,' the purpose of which is to determine the functional characteristics of X- and Y- carrying sperm cells so that one pool of sperm gains an advantage over others during the movement towards the egg-cell, and another pool of sperm does not gain such an advantage. Clearly, this is a universal mammalian biological mechanism which is regulated by hormones, and can be the subject of external manipulation. Note that the above-defined cyto-genetic phenomenon occurs in the male organism. In this connection, we should turn our attention to the sexual dimorphism of spermatogenesis and ovogenesis. 5. Sexual dimorphism of spermatogenesis and ovogenesisThe two sexes differ fundamentally in gametogenesis. The basic difference is that spermatogenesis is a dynamic, strictly ordered process of forming new cells, whereas in a woman an already formed cell is induced to develop further. The male organism, therefore, can be influenced by outside forces, and the female can transfer these influences to successive generations.
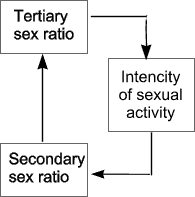
The sexual dimorphism of gametogenesis allows for the realization of the so-called 'population' functions of gender 6. Population functions of genderDifferences in gametogenesis are necessary in nature to ensure the realization of the population functions of the sexes. According to our current understanding, the female sex represents 'long-term memory' and the male 'operating memory' (25, c.108). This idea helps to understand the phenomenon by which the level of stress in a population influences the rate of male births. The idea of the population functions explains the difference in average life expectancy for males and females. More accurately, these differences are a consequence of the population functions. A difference in the average life expectancy of males and females indicates a critical situation for a population, especially in cases where male life-expectancy is on the decline. This indicates that the population is under the influence of negative factors. The influence of these factors is even more apparent if the rate of male births increases. The secondary sex ratio and the difference in average life expectancy provide the clearest evidence for the population functions of gender. The functions have an adaptive purpose and are responsible for the continued existence of a population and its adaptation to the environment. Well known in the field of demographic statistics is the so-called 'war phenomenon,' whereby the rate of male births rises in the years during and after a war. One of the proposed explanations for this is the natural increase in marriages after wars and other social upheavals. Clearly the ratio of the sexes is dependant on specific human behavior. It is well established that in the normal course of the reproductive process, neither male nor female embryos are selectively eliminated (the primary sex ration does not change). Therefore 'the change in the secondary sex ratio in favor of male births is conditioned by the selective fertilization of eggs by Y-carrying sperm' (28, c.38). The sex ratio of a population is a typological characteristic, usually characterized by stability. Under the influence of extreme factors, however, it can shift significantly, returning to its normal level only after the disappearance of these factors. All this is evidence in favor of the theory that the mechanism for regulating the sex ratio is consistent and non-random. The hypothesis that males and females are conceived with equal probability was refuted long ago. The intensity of sexual life plays an important role in the mechanism of sex determination. 7. The Mechanism of negative reverse connection in sex ratio regulationMore than 30 years ago, specialists postulated that intensity of sexual activity is a mechanism of negative reverse linking between the secondary and tertiary ratios of a population. This is easy to explain. When there are more males than females in a population, then the rate of male sexual activity decreases, and conversely, when there are more females then males, it rises. All this follows common logic, and does not require further explanation. From this insight, we can draw some conclusions. In the case of a preponderance of males, the rate of female births should rise, so that in the next generation the sexes will balance out. In the case of a preponderance of females, the male birth rate will rise, and the sexes will also balance out.
Diagram 3. The regulation of tertiary and secondary sex ratio through the intensity of sexual activity
Diagram 4. Sex hormones in man during increasing sexual activity rhythm Since the general intensity of sexual activity in a population is an aggregate of the sexual lives of many individuals, the study of individuals should be of particular interest, all the more so considering that significant variability between individual sexual behavior has already been established. In men, the rhythm of sexual life is determined by the specifics of the system of sex-hormones, and the influence of the above-mentioned cyto-genetic mechanism of pre-zygotic determination, including glandulocyte, a link in the process of spermatogenesis at the stage of the transformation of spermatocyte-1 to spermatocyte-2, and the mechanism of crossing-over. 8. Variations in the rhythm of sexual activityWe must turn our attention to the condition of the human organism at various levels of sexual activity. It has been established that with frequent ejaculations, several of a man's sperm indexes change.The fertilizing abilities of the sperm are not, however, reduced. Some well known studies have shown that the incidence of infertility in prostitutes is linked to high levels of spermicidal anti-bodies in their blood serum. 9. The importance of the frequency of male ejaculationIt is well known that periods of sexual abstinence encourage the conception of female children, and that frequent sexual activity leads to the conception of more males. To quantify the categories of 'frequent' and 'infrequent' we use the theory of sexual constitution. We understand sexual constitution as the aggregate of stable biological mechanisms, arranged under the influence of hereditary factors, and conditioned during the pre-natal period and in early stages of ontogenesis. Sexual constitution fixes the individual sex-drive at a certain level, and determines a subject's resistance to pathogenic factors which have a selective role in the sexual sphere. Using the algorithm for determining sexual constitution (by G.Vassiltchenko), we can calculate the genotype index (Kg), the index of sexual activity (Ka), and also their ratio (Ka/Kg). The ratio of the two indexes allows us to quantify the rhythm of ejaculation as shown below: 1. Ka/Kg between 0.90 - 1.09 is evaluated as an "optimal" rate of sexual activity. 2. Ka/Kg higher than is evaluated as "over-active" sexual life. 3. Ka/Kg lower than 0.89 is evaluated as "relative abstinence". The use of the algorithm of sexual constitution, then, helps to evaluate and to classify the rate of sexual activity. In general, the rhythm of sexual life is a product of libido, and it is worthwhile to examine the causes of variations in the level of sexual activity. 10. Ideas about the relationship of the level of sexual desire to the rate of sexual activity
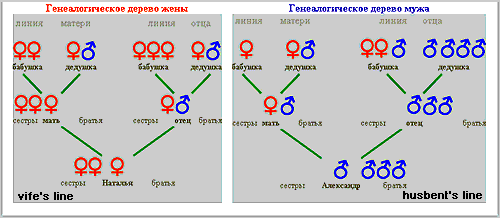
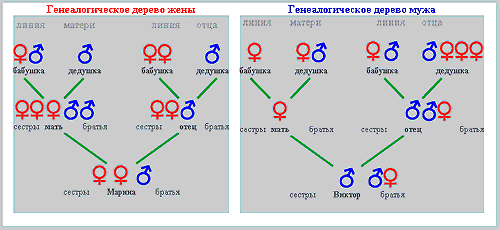
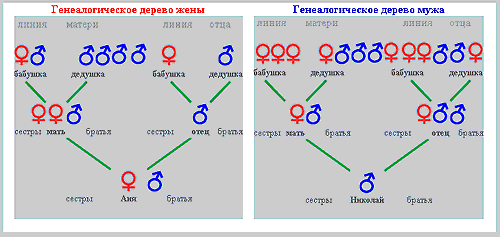
In the first state the rate of sexual activity corresponds to the level of libido, in the second case it is higher, and in the third it is lower. Continuing the theme, let's take the example of a man whose basic level of libido is classified as 'low,' but who is driven by personal factors towards an intense rhythm of sexual life - a result of his acceptance of the social stereotype that in the first year of marriage sexual intercourse should be "frequent". As a result of this man's high rate of sexual activity, the cyto-genetic mechanism of pre-zygotic determination produces an elevated level of testosterone, and the pool of Y-carrying spermatozoids have a selective advantage, which is realized in the process of pre-zygotic selection, leading to the conception of a male child. There are some families where the genealogical tree along the male line shows only men (and vice versa) for several generations. >>> It is possible that in these cases, the limited level of sexual desire is inherited, as are the specific personal factors which lead to "frequent" sexual activity. We will now examine the second case - where the man has a 'low' or 'moderate' level of libido, but the personal factors are situation-dependant, and not genetically determined. The man's partner has a strong sexual drive, and the couple's sexual life reflects this. The man again produces high levels of testosterone, leading again, through the familiar process, to the conception of a male child. In this case, the subject's genealogical tree does not have the characteristics described above. In the next example, the genetically determined level of libido is 'high,' but personal and social factors (for example, a religious world-view) limit the level of sexual activity. As a result the cyto-genetic mechanism produces less testosterone, and the X-carrying pool of sperm gains a selective advantage, which is realized in the process of pre-zygotic selection, leading to the conception of a female child. We will not continue to enumerate all the possible variants, but note merely that, even though the factors influencing the basic level of libido and the rate of sexual activity are extremely numerous and varied (these factors include the influence of the sexual partner, somatic condition, marital status, social conditions, and so on), the modulations of the cyto-genetic mechanisms of pre-zygotic determination lie within certain clear boundaries. Yours sincerely, Yuriy Zharkov 11. Clients' genealogical trees
Picture 1: Genealogical tree of couple A-N
Picture 2: Genealogical tree of couple V-M
Picture 3: Genealogical tree of couple N-A |
|||||||||||||||||||||