A CRITICAL EVALUATION
|
|
Department of Anatomy, University of Louisville School of Medicine, Louisvile
Currently at: University of Hawai`i at Manoa John A. Burns School of Medicine Department of Anatomy & Reproductive Biology Pacific Center for Sex and Society Honolulu, Hawai`i 96822 <diamond@hawaii.edu> Phone: (808) 956-7400 Fax: (808) 956-9481 Published in The Quarterly Review of Biology Volume 40, No. 2, June 1965 |
ABSTRACT |
|
|
The classical view of human sexuality holds that man is invested with a particular sex within which he, as an individual, develops. Recent attempts to alter this conception and to explain psychosexual maturation as developing from a neutral rather than a sexual base are here reviewed and criticized. Essentially, a psychosexual neutrality-at-birth theory holds that male and female patterns of sexual orientation and behavior are attributable exclusively to learning or imprinting phenomena. This theory is derived from clinical observations of individuals manifesting morphological sexual incongruities (hermaphrodites, pseudohermaphrodites, etc.). This article defends the view of inherent somatic sexuality organizing man's psychosexual development by: (a) reviewing man's place on the evolutionary continuum, and the broad base of sexual behavior within which this discussion must be considered; (b) presenting normative, clinical and anthropological evidence inferring a particular sexual predilection at birth; (c) showing genetic, hormonal. and neural indications for sexual predisposition; (d) refuting the extent of imprinting involved in humans; and (e) showing the futility of separating "nurture" from "nature" in reference to the role of learning and acquisition of a gender role. |
|
INTRODUCTIONA review of the many areas pertinent to the field of sexual behavior over the last decade reveals the development and elaboration of various psychosexual medical aspects. This in itself is not surprising in view of the greater all-round interest in the psychological field coupled with the increased publicity given to studies bearing on sexual behavior. What is surprising, however, is a certain direction this development has taken and the relative ease with which one view has been accepted. Starting in 1955, articles written by John Money and Joan and John Hampson, either in collaboration or separately, began to appear with regularity. Within two years these investigators had produced a book and almost a dozen papers (see list of literature). The content of their articles details clinical examinations, descriptions, interviews, and therapy of various individuals with sexual abnormalities. Particular attention was given to those patients classified as sexually precocious or as hermaphroditic. |
|
|
The term "hermaphroditic" is broadly used here to indicate sexual deviance from the normal condition in any two or more of the following ways: (a) external genital morphology; (b) internal accessory reproductive structures; (c) hormonal sex and secondary sexual characteristics; (d) gonadal sex: and (e) chromosomal sex. |
|
|
This work must be considered of value in giving new insight into various clinical areas previously almost taboo, and in shedding light on some particularly intriguing questions of human sexuality. The articles went further, however, into theorization about the ontogeny of human sexual behavior and its modifiability, and included a reappraisal of classical notions of sex roles. Probably on the strength of the clinical aspects, this revised theory seems to have gained favor and gone without serious challenge. Essentially the theory advocated by Money and the Hampsons holds that gender role - all those things that a person says or does to disclose himself or herself as having the status of boy or man, girl or woman. respectively, and sexual orientation as a male or female - is independent of chromosomal sex, gonadal sex, genital morphology, hormonal balance, or other commonly used indicators of sex (Hampton and Hampson, 1961; J. L. Hampson, 1964). In their own words: |
|
|
. . . in place of a theory of instinctive masculinity or femininity which is innate, the evidence of hermaphroditism lends support to a conception that psychologically, sexuality is undifferentiated at birth and that it becomes differentiated as masculine or feminine in the course of the various experiences of growing up (Money, Hampson, and Hampson. 1955b). Now it becomes necessary to allow that erotic outlook and orientation is an autonomous psychologic phenomenon independent of genes and hormones, and moreover, a permanent and ineradicable one as well (Money, 1961e). It is more reasonable to suppose simply that, like hermaphrodites, all the human race follow the same pattern, namely, of psychosexual undifferentiation at birth (Money, 1963a). Thus, in the place of the theory of an innate, constitutional psychologic bisexuality . . . we must substitute a concept of psychologic sexual neutrality in humans at birth (Hampson and Hampson, 1961). |
|
|
In brief. their theory may be called a psychosexual "neutrality-at-birth" theory, as opposed to a "sexuality-at-birth" theory. Although other investigators have supported and projected evidence and theory of a similar nature, the present focus is placed on the works originating from the authors just cited, since they, starting almost with a formal challenge to the classical concept of human sexuality (Money Hampson, and Hampson, 1955b), are perhaps at present most closely associated with this approach and most prolific and influential it this area of thought, particularly in regard to clinical considerations. It is my present intention to review the evidence relative to this theory. and to suggest in contradistinction that the very same data may not be inconsistent with more classical notions of inherent sexuality at birth. This inherent sexuality, like other biological characters, need not necessarily manifest itself at birth as it might be first revealed at puberty or during adulthood. Nevertheless, inherent sexuality may, from birth, provide a built-in "bias" with which the individual interacts with his environment. Generally the concept of psychosexual neutrality at birth may be considered to draw support from the following three broad areas: (1) clinical cases; (2) the imprinting phenomenon; and (3) learning theory. The clinical material will be considered first, as it has figured most prominently in the formation and presentation of this theory. Wherever possible, counter-evidence will be drawn primarily from human data. CLINICAL EVIDENCE IN SUPPORT
|
|
|
In the light of hermaphroditic evidence. it is no longer possible to attribute psychologic maleness or femaleness to chromosomal, gonadal or hormonal origins, nor to morphological sex differences of either the internal accessory reproductive organs or the external genitalia. . . From the sum total of hermaphroditic evidence the conclusion that emerges is that sexual behavior and orientation as male or female does not have an innate, instinctive basis. In place of a theory of instinctive masculinity or femininity which is innate, the evidence of hermaphroditism lends support to a conception that psychologically, sexuality is undifferentiated at birth and that it becomes differentiated as masculine or feminine in the course of the various experiences of growing up. |
|
|
During these experiences of growing up the normal human is assumed [from hermaphroditic (pathological) data] to be imprinted as well as taught a sexual role. The first 2 1/2 or 3 years of life are supposedly the critical period for human imprinting, and cases are cited where alteration of sex after this age is traumatic (Money, Hampson, and Hampson, 1955a, 1956, 1957: Hampson and Hampson, 1961; Money, 1961e). This age is correlated by Money with the development of verbalization. He believes that the critical period for the imprinting of gender role and orientation corresponds with a critical period for the establishment of a native language. Another critical period for limited modification of gender imprints is postulated to occur at puberty (Money, 1961e). To further the idea that a psychosexual imprint is fixed and irreversible, 5 cases of genetic females born with fused labia and enlarged clitorides are cited. Two of these were reared as boys and three as girls, and yet all seemed to be psychologically content in their present sex (Hampson and Hampson, 1961). The extent and depth of the imprint and subsequent learning is supposedly such that alteration after 4 years of age is fraught with psychological danger and is usually unsuccessful (J. G. Hampson, 1955, 1964). "With only I exception the 6 patients reassigned later than the first birthday were rated as inadequately adjusted" (Hampson and Hampson, 1961). Given the evidence presented, what may we conclude? It has been shown that hermaphroditic individuals in our society find it possible to assume sexual roles opposite to their genetic sex, morphological sex, etc., and they can assume this role so well that they can function socially as "normal" members of society, engage in erotic activities. and receive pleasure in their reared roles. To best assume this role it is most advantageous in our society for them to start early in life. preferably before the first birthday. Beyond these conclusions, however,much has been extrapolated. Maybe it has been strongly demonstrated that humans, particularly hermaphroditic ones, are flexible when it comes to the assumption of an incongruous sex role. Yet to assume that a sex role normally is exclusively or even mainly a very elaborate, culturally fostered deception and imprinting phenomenon, and that it is not also reinforced by taboos and potent defense mechanisms superimposed on a biological prepotency or prenatal organization and potentiation seems unjustified and, from the present data, unsubstantiated. CONSIDERATIONS REGARDING INHERENT
|
|
|
Because they are often confused and misused, the terms "innate" and "instinct" are defined for use in this paper as follows: "Innate, a term applied to differences in genetic character between two members of the same species that have been raised in the same environment. (It is now generally acknowledged that the term innate, a technical one in genetics, cannot properly be applied as synonymous with unlearned or inborn. and its use in that sense may be expected to become less frequent- Where it has appeared in the past, innate behavior should now be read as unlearned behavior or species-specific behavior)." (Verplank, 1957). "Instinct, a hierarchically organized nervous mechanism which is susceptible to certain priming, releasing, and directing impulses of internal as well as external origin, and which responds to these impulses by coordinated movements that contribute to the maintenance of the individual and species." (Tinbergen, 1951). |
|
|
I will hold that man in regard to his sexual behavior patterns is, like all other vertebrates, subject to prenatally organized mediation. The manner or extent of this mediation is not yet clear but is believed to involve the fetal organization and potentiation of certain neural tissues which are, within genetic limits, postnatally modifiable but not to the extent of complete reversal or negation. This effect on the nervous system is believed primarily to be a function of the genetically induced endocrine environment of the presexually differentiated individual. For man as well as most other vertebrates this is a prenatal occurrence. Ontogenetic experiences are superimposed on this potentiated nervous system and serve to give emphasis and further direction to predisposed tendencies. Man is probably more flexible in regard to this organization than any other species, but that would not justify our saying he is free of it. (2) It is significant that no criteria or definition of human male or female behavior has found universal acceptability. Humans, as well as many other species of animal, normally exhibit elements of sexual behavior usually attributed to members of the opposite sex. The capability and frequency of such behavior is neither rare nor bizarre. I will defend the point, that although humans can adjust to an erroneously imposed gender role, (a) it does not mean that prenatal factors are not normally influential, and (b) they do so with difficulty if not prenatally and biologically predisposed. Beach in a lecture given at Yale in 1948 pointed out that: |
|
|
. . . laws are violated by the homosexual individual but to describe his behavior as `unnatural' is to depart from strict accuracy. The zoological evidence shows that female mammals frequently display masculine coital behavior when confronted with sexually receptive members of their own sex. This has been observed in more than a dozen species and undoubtedly occurs in many others not yet studied. . . . The physiological mechanisms for feminine sexual behavior are found in all males and those for masculine behavior exist in all females. The same stimuli that elicit feminine copulatory reactions in the female will, under appropriate conditions, produce similar reactions in many males; and the stimulus configuration evoking masculine responses in males is the one which most effectively calls forth these same responses on the part of the female. Human homosexuality reflects the essential bisexual character of our mammalian inheritance. The extreme modifiability of man's sex life makes possible the conversion of this essential bisexuality into a form of unisexuality with the result that a member of the same sex eventually becomes the only acceptable stimulus to arousal . . . . Human sexual life is not unique in its susceptibility to modification . . . . (Beach, 1948a [emphasis added.]). |
|
|
Kinsey and colleagues, indeed, report various varieties of heterosexual and homosexual activities as not uncommon either for males or for females (Kinsey, Pomeroy, and Martin, 1948; Kinsey, Pomeroy, Martin, and Gebhard, 1953). Thus, both normal human beings or hermaphrodites who exhibit various so-called anomalous sexual behavior still are performing within the biological continuum predictable by evolution. Evolving from a highly stereotyped pattern among primitive organisms, humans are capable of displaying highly flexible sexual behavior patterns. An evolutionary trend starting with inflexible stereotyped sexual behavior and progressing to flexibility in behavior is consistent with modern genetic and evolutionary concepts (Tax and Callender, 1960). The very considerable extent of this flexibility, particularly in hermaphrodites, may account for the many cases of maintained gender role which are incompatible with morphological criteria of sex. This same flexibility may account for the erroneous theory of psychosexual neutrality for normal individuals. EVIDENCE IN SUPPORT OF SEXUALITY AT BIRTHThe evidence presented in support of the concept of psychosexual neutrality-at-birth primarily involves hermaphroditic individuals who have successfully adapted themselves to an assigned gender role inconsistent with one or more morphological criteria of sex. Once reared in a malassigned sex past the age of four, these individuals are supposedly unable to negotiate a change in their gender role without severe emotional trauma (Hampson and Hampson, 1961). The neutrality theory is supported by no normative data. A theory that psychosexuality exists at birth, on the other hand, can use the same evidence and demonstrate (a) that humans, hermaphrodites in particular, are flexible enough to maintain an atypical gender role, either by choice or accident, even when contradictory to their normal external genitalia; (b) that a gender role, if malassigned may be reassigned after the age of four without undue trauma; and (c) irrespective of the foregoing conclusions, that normal as well as hermaphroditic individuals are predisposed toward a particular gender role at birth. Studies of Normal ChildrenAlthough the literature is scanty in this regard, various studies of neonatal, preverbal, and preschool children indicate that there are differences which may be considered indicative of sexual differences existing from birth and which are psychosexually predisposing to adult male or female roles. Terman (1946), in an extensive review of normal psychological sex differences in children, wrote: "Sex differences have been found for almost every physical variable, including body build, gross and fine anatomy, physiological functioning and biochemical composition. Indeed, every cell in a human body bears the stamp of its sex." He then proposed that some of these differences, such as height, weight, body build, strength, endurance, motor ability, and rate of maturation, might be expected to reflect themselves behaviorally. In this regard he lists the following: (a) Body Size The mean weight of boys at birth exceeds that of girls by approximately 5 per cent and their body length is greater by 1 or 2 per cent. This superiority continues until about the age of 11. (b) Vital Capacity. As one of the determiners of the sustained energy output which is possible for an individual, vital capacity may considerably influence sex differences in play interests, and other activities. Boys show a 7 per cent superiority in vital capacity by age 6 and rapidly increase their lead to about a 35 per cent superiority at age 20. (c) Muscular Strength. Tests of strength in grip, back, and legs show boys to be superior at all ages tested. At age 7 boys show about 10 per cent superiority and this increases to a 50 to 60 per cent advantage at age 18. (d) Rate of Maturation. It is widely recognized that girls generally exhibit more rapid physical development than boys. In skeletal development girls are superior to boys at birth and increase this superiority at a fairly steady rate until growth is complete. "Girls of all races precede boys on the average by twelve to twenty months in pubertal development, and their adolescent growth changes are correspondingly accelerated." These obvious examples of constitutional differences are reflected in the individual choice of the more muscular forms of behavior of male children, such as running, climbing, and wrestling, compared with the more conservative and sedentary activities of female children, such as hopping and skipping. This was extensively reported by Lehman and Witty, as early as 1927, in a classical study involving some 19,000 children. Walker (1962), studying the somatotypes and behavior of preschool children 2 to 4 years old, has presented much data to indicate that there is a good correlation between sex-related behavior traits and body build as rated by endomorphy, mesomorphy, and ectomorphy. He concludes that "in this group of preschool children, important associations do exist between individuals' physiques and particular behavior characteristics. Further, these associations show considerable similarity to those described by Sheldon for college-aged men, though the strength of association is not as strong as he reports. It is suggested that the relations are multiple determined, arising from primary body conditions (e.g., strength, energy, sensory thresholds) . . . . In particular, variations in physical energy, in bodily effectiveness for assertive or dominating behavior. and in bodily sensitivity appear as important mediating links between physique structure and general behavior." In areas such as conation and cognition we also note early manifest sex differences. This is especially significant, since these categories are believed to reflect traits less influenced by culture. There is considerable evidence that from early childhood boys show more aggression and anger than girls. Goodenough (1931) reported this to be manifest as early as 7 months of age. In regard to personality Walters, Pearce, and Dahms (1957) have demonstrated that 2- to 5-year-old girls are significantly more friendly, sympathetic, and helpful to others ("affectionate") than are boys of the same age; while boys tend to display more actual or threatened hostility to others ("aggressive"). Walker (1962) found similar results for children of the same age range and in fact remarks that the difference is as great at 2 years of age as later. Boys and girls are seen to show different perceptual responses to Rorschach forms (Ames, Learned, Métraux. and Walker, 1952). They have shown, for example, that throughout the first ten years of life boys give more responses than do girls in every category. Girls respond differently to color than do boys, and they respond more often to the perceived form alone. The type of response also seems sex-dependent. At 3 1/2 years of age boys give more responses than do girls and verbalize more. Boys see more movement in the figures, and boys make more mention of urogenital structures. At 7 years of age sex differences in perceptual responses are the most marked. Girls' responses tend to be neater and more concise. Boys' records are longer and more involved, their comments and explanations complicated and rambling. Girls give more global responses, boys more details and tiny details. Boys give more aggressive responses. Ames and Learned (1954) showed significant sex differences in developmental trends in block building and block construction. Like the Rorschach test, this block-building and construction test supposedly reveals inherent differences and forms of behavior not significantly influenced by culture. Ames and Learned used 350 children equally divided between the sexes at ages 2 through 6 years. Conspicuous sex differences occurred at all ages tested For example, at 2 years "boys build in a more complex and detailed manner than do girls." Boys show more responses and show more compact designs. At 4 years, "The structures of girls fall more than those of boys. Twice as many boys as girls make one large compact structure, and many more boys than girls make scenes." At 6 years: "Nearly twice as many boys as girls build symmetrically. Many more boys than girls build one large compact structure. Girls show more interest in size and color. More girls than boys purposely destroy their products." Many more normative studies of similar character could be presented. In effect, the conclusions and implications of their findings are summarized by the pediatrician Benjamin Spock. Spock (1964), in an article entitled "Are we minimizing differences between the sexes?" lists sex differences noticeable from birth in areas of interest, aptitude, and personality, and concludes: "In terms of basic temperament and drive there are fairly consistent differences between the sexes, though they may be accentuated or obscured by upbringing." He then prescribes that for greater security in adult roles it is better to capitalize on these differences rather than to minimize them. The following section presents cases where these inherent differences were frustrated by improper assignment of sex role. Flexibility and sex reassignmentClinical Evidence. Dicks and Childers (1934) presented a case of a genetic male child with hypospadias who from birth to the age of 14 was treated and dressed in every way as a girl. As the individual developed, however, he began to doubt his sex assignment and his interests became "as nearly those of a healthy adolescent boy as would be possible under the circumstances." Despite the insistence of his parents, he had no doubt as to his sex and urgently demanded a transformation to his proper masculine role. After a change was initiated no conflict regarding sexual matters was apparent, and "his social adjustment was so phenomenal that at no time was there indicated a need for intensive psychotherapy." Armstrong (1955) has reported a case of a male pseudohermaphrodite, a male with undescended testes and hypospadias, who lived as a girl for 13 years. At the age of 13 years this condition was revealed, and a plastic repair of the genitalia was performed. The patient subsequently assumed the male sex very successfully. In reviewing cases of this sort Armstrong analyzed various problems inherent in this treatment but said. "It should be our duty as medical practitioners to repair physical abnormality and advise the patient to assume his or her true sex." Ghabrial and Girgis (1962) reported two cases of males who were reared from birth as females and lived as females until, when old enough to rebel successfully, declined to continue their malassigned roles. The first individual changed to a male role at the age of 14, and when seen a year later seemed cheerful, happy, and apparently well adjusted. The second underwent this change of role at the age of 20, despite a penile amputation at birth (it being confused for a hypertrophied clitoris). Notwithstanding the many years and opportunity for gender role learning, rehearsal, and imprinting, both patients insisted on sex reassignment to agree with the sex to which they felt they belonged. Norris and Keetel (1962) presented a case of a female with "a congenital anomaly of the vagina" who shortly after birth was diagnosed as having a bifid scrotum with infantile penis. The child was raised as a male, with corrective surgery to be done at a later date. At puberty the mistaken diagnosis was revealed and the patient decided, upon advice from the physicians, to become a female. Plastic repair of the genitalia was done at age 17. At age 20 a psychiatric examination revealed "no apparent neurotic modes of adjustment, no distortion of personality; she related spontaneously and with warmth . . . . a healthy individual: no indices of a neurotic or psychotic mode of adjustment were elicited." She was then happily married with a full and complete sexual adjustment. In regard to general comments in the area of role assumption, Norris (personal communication) says: |
|
|
I feel, as a psychiatrist, that people are amazingly flexible in their ability to handle roles. This is evident in many other aspects of living. Immigrants come to a new country where they must speak a new language and adopt a new social and vocational role, and most of them adapt to this well. Men are taught the harm of aggression and taught not to kill, but during wars are able to adapt to the concept of doing so after a few months training, and furthermore, after the war is over, most return to civilian life with relatively little difficulty, and certainly here are role changes occurring at a very late age, which stable people handle without too much trauma. I don't believe that sexual role changes should be any more difficult, assuming an intact personality. |
|
|
Dewhurst and Gordon (1963), in an extensive survey of 18 patients in whom a correction of malassigned sex role was made after the age of 4 1/2 years, contend that 15 of the individuals involved assumed the new role with considerable success. They correlate this with the inherent feelings of many of the individuals concerned that they were being reared in the wrong sex. These findings are related to the views of Money and colleagues and are considered as non-supporting and contradictory. Dewhurst and Gordon (1963) write: |
|
|
. . . several parents, including one couple who had badly wanted a child of the sex in which theirs was being reared, believed that they had observed behaviour more appropriate to the opposite sex. Arguably, if uncertainty had not been expressed, these views might not have been held, but we are inclined to reject this, preferring to believe that the child's natural tendency to the gonadal sex will sometimes militate against the sex of rearing and may even overcome it. |
|
|
The following two clinical reports may be considered as crucial for the theories involved. The first case involves a rare patient who, as an unambiguous male was raised from birth as a female (Dr. Robert Stoller, personal communication). If gender role is independent of constitutional factors and is dependent only on sex of rearing and learning, this individual should have developed psychosexually into a female. Not only was this not seen, but in this normal individual rearing seemed to have very little effect. Stoller writes: "Despite attempts by the parents to make this child a girl, almost from birth on the child refused to be comfortable in the assigned sex or sex of rearing, continuously fighting all attempts from her feminine mother to be a feminine daughter. At the age of 14 we found her to be a genetically and endocrinologically normal male. The patient shifted overnight to a completely normal boy -a most remarkable and successful change." The second report involves an individual who was raised as a girl and "appeared to be a girt insofar as social status and outward sex status was concerned" (Brown and Fryer, 1957). Despite a female sex assignment without ambivalence and the body habitus to match, the patients "normal feelings and reactions" in themselves were strong enough to prompt a medical consultation concerning the true sexual identity (Brown and Fryer, 1957, 1964). Clinical examination confirmed the suspicion of mistaken identity and "the patient immediately recast his life," and received corrective surgery. A follow-up study after 10 years showed the patient happily married and a proud father (Brown and Fryer, 1964). More cases could be cited. In effect they serve to demonstrate that inherent male or female sexual orientation, feelings, and disposition may develop despite body habitus, sex of assignment, or rearing. They also indicate that if an incorrect assignment is made, a change in role may be possible without pertinent psychological trauma. It may be noted that most of the cases involved individuals at puberty. This is a time when inherent diasthetic sexuality, now spurred by maturation, would indeed be expected to manifest itself most strongly even if contrary to a malassigned role. This is what might be predicted from a theory of inherent sexuality, yet could not be predicted from a theory of sexual neutrality. It might be asked from what do these doubts stem, since the family and society are reacting to the individual as a member of the assigned sex and it is within that capacity that reinforcement is found and outside of that capacity where difficulty would be encountered. To explain these nontraumatic sex corrections on the basis of a theory of inherent psychosexual predisposition is no problem. The individual is settling into a role for which he or she is more suited constitutionally, and may even feel relief in the new role. On the other hand, to explain not only the origin of doubt as to proper sex assignment but the ease of sex correction on the basis of a theory of psychosexual neutrality would necessitate postulating for the initial gender assignment incomplete imprinting or misprinting, faulty learning, other hypothetical conditions, or a combination of these, none of which can be adequately substantiated. This is especially clear in the cases presented, since it would be difficult to imagine the subsequent imprinting or learning necessary for the new sex role to be more effective than that for the initial sex role. It would be theoretically more parsimonious to accept the view that inherent in each individual is a diasthetic predisposition to a particular gender role spectrum. If placed in a situation outside of this particular spectrum, the individual finds difficulty in adjustment. Anthropological Considerations. Among societies in which mores permit such changes, an individual may voluntarily assume a gender role inconsistent with the one in which he has been reared and an individual may alter his gender role well after the so-called "critical period;" all in keeping with accepted modes of their respective societies and thus presumably maintaining psychological normalcy. Cultures also exist in which, even though gender roles are not strictly assigned, the development of sexual behavior follows a normal nonambivalent course. These societies may thus provide evidence as to the relative roles of heredity and environment in sex role status. In societies of the first type the sex of rearing may exert little evident effect upon the individual's adult choice of sex role, and in societies of the second type proper sexual orientation occurs despite the lack of strict role assignment. These situations would seem to indicate an inherent predisposition that functions despite the presence or absence of strong environmental ontogenetic influences. Some of the American Indians provide an example of a society where sex role changes are permitted despite strong orientation during rearing (Mead, 1961). Mead says that in tribes where a berdache [transvestite male] was recognized as a possible sex career, "male children were watched and tested from an early age - were they going to be 'braves' or 'live like women'? Once the choice was made, elaborate prescriptions of correct social behavior were available" (emphasis added). The two points to be made here are the following: (1) that the determiners of the gender role to be assigned were not based on morphological sex but on innate and on instinctive self-elaborated behavioral manifestations of the individual; and (2) that, without trauma to the individual, this role can be assigned and accepted by society postpubertally. Mead (1961) gives an illustrative example: "At the time when his bodily candidacy was remarked there was no living berdache in the tribe, but the women began watching this boy, and once undressed him to see if he, whose behavior appeared to them as feminine, 'really was a male.' Having satisfied themselves as to his external sex morphology, they then pronounced him to be a berdache. He wore male exterior clothing but female underwear, was unmarried . . ." (emphasis original). Thus the berdache is a recognized recurrent sex role that seems to arise independently in an individual who possesses unambiguous male genitalia and is raised in all ways as a male. These individuals, despite their rearing, begin to manifest feminine behavior patterns and are assigned to a sex role in accordance with this inconsistency. Corroborating the assertion that gender role assignment may be based on an individual's behavior rather than be a determiner of it are the various criteria that have been established among some ethnic groups to aid in determining just such an assignment. Bravery is the determining masculinizing point among Plains Indians (males being considered and treated as females unless brave) (Mead, 1961), and preference for womanly occupations is the female criterion for males among Samoans (males being considered and treated as females if they prefer womanly occupations) (Mead, 1928). Other anthropological evidence can illustrate the development of male and female sexual patterns in spite of the lack of specificity in rearing. Data are available for communities where there is no sex gender in the language and the names of males and females are undifferentiated and for communities where juveniles of both sexes dress alike (Mead, 1961). Societies exist where boys are classified with women until initiation (Iatmul) and where girls are classed with men until betrothal (Manus) (Mead, 1961). Here it would seem that the critical period would be passed without opportunity for gender role imprinting and learning! To think or to expect that these individuals would exhibit no adult sexual behavioral distinctions or that they would remain in a sort of sexual limbo is not in keeping with the facts. It no doubt would aid in the sexual orientation of individuals within these cultures if their society had strict gender taboos and restrictions. But in their absence, constitutional factors as well as obvious physical characteristics are sustaining. Notably, what evolves from anthropological studies is scattered but abundant evidence that there occur individuals with the behavior patterns of the opposite sex, in the absence of any patterned cultural recognition of such a possibility. This conclusion strongly suggests the presence of some rare but recurring constitutional factor which is less overt than congenital genital anomalies. It seems safe to assume that any behavior, sexual or not, which can be institutionalized in a culture and regarded as a recurrent possible human choice has some hereditary, i.e., genetic, basis (Fletcher, 1957: Mead. 1961). Fletcher (1957), in defending the role of sexual instincts in man, also calls upon cross-cultural data. In concluding this portion of his discussion he criticizes the attempt to explain all human behavior in terms of learning cultural direction and values. His argument is noteworthy: |
|
|
Two points which have a bearing on the present question might be mentioned. Firstly, it seems probable that the tendency to describe individual personalities in terms of broad cultural uniformities, has gone too far, and it may be that the whole question of the degree to which individual persons are 'conditioned' by their 'culture' has been too readily assumed, rather than demonstrated. The second point is that the emphasis on the varieties of behaviour in different human societies may well have tended to obscure the similarities; similarities not only of 'instinct-experience' (congenital impulses), but even of elements of behaviour. Thus, let us consider behaviour involved in walking, eating, drinking, defecation, sleeping and waking, sexual intercourse, the care of young infants, fear and the avoidance of danger. It would seem that, no matter how varied are the more complicated methods of food-seeking, sanitation, provision for rest and sleeping, the regulation of the relations between the sexes, the bringing up of children, the social methods for avoiding danger, and so on, it remains true nonetheless, that there are certain basic features of behaviour which are congenital in man and which are necessarily involved in the satisfaction of the congenital cravings, some of which persist throughout his life. Margaret Mead describes for us three varieties of sexual behaviour in three societies, but, we suggest, this merely indicates that the cultural setting exercises its influence upon the behaviour adopted in order to satisfy the instinctual demand which is inherited: and the contributors to the theory of instincts would have no quarrel with this. But if the persisting sexual impulse was not inherited, and if this impulse did not drive men and women to definite fundamental forms of behaviour for its gratification, no 'social conditioning' would ever take place. There would be nothing to 'condition.' No rules regulating sexual relations would be necessary. There would be nothing to regulate. If the Cultural Anthropologists [or any other group] are going to make us doubt the existence of the sexual instinct in man (and the same applies to the other instincts) they must show us not how variously men and women satisfy this instinct in different societies, but a society in which-as an outcome of the determining influence of the social environment - there is no evidence of the sexual impulse, or of the basic forms of sexual behaviour, at all. We can predict that such a society is going to be very hard to find. |
|
Predisposition of Gender OrientationGenetic Considerations. Primary in any discussion of predisposition towards or away from a particular pattern of sexual behavior would be consideration of the possible genetics involved. That animals are subject to inherited tendencies in relation to their sex roles cannot be disputed. Young (1957) has reviewed many of the nonclinical studies and presents evidence not only showing innate manifestations of sexual behavior patterns but describing how different animal strains can react differently to the environment. Many recent studies corroborate these findings (Goy and Jakway, 1959; Jakway, 1959: McGill, 1962a, b; McGill and Blight, 1963a, b). In regard to man, however, our knowledge is a little more obscure. Kinsey, Pomeroy, and Martin (1948) state this theme thus: "The most important biologic factors affecting the nature and frequency of sexual response in the human animal are the hereditary forces which account for the differences between male and female. Within either of these sexes, heredity must also account for some of the variation in sensory structures and in the mechanisms which are concerned with emotional response." However, no other area in the study of human sexuality seems beset with as many difficulties as a study of inherited sexuality. Not only would a proper investigation involve a broad longitudinal survey over many years but it would have to cover succeeding generations. Nevertheless some data are available. Kallmann's work (1952a. b) and that of Schlegel (1962) are probably the most conclusive. Kallmann studied 40 monozygotic and 45 dizygotic twin pairs in which one of the cotwins was a known overt homosexual. He found 100 per cent of the monozygotic twins concordant for homosexuality; whereas the dizygotic cotwins were essentially similar to the general male population. Strikingly, the monozygotic twins were even comparable in the mode and extent of their deviance and type of displayed behavior, and although they had developed their gender roles independently when separated from each other. Kallmann considers this evidence, and I would agree, to throw "considerable doubt upon the validity of purely psychodynamic theories of predominantly or exclusively homosexual behavior patterns in adulthood and correspondingly strengthens the hypothesis of a genically determined disarrangement in the balance between male and female maturational (hormonal) tendencies" (Kallmann, 1952a). Schlegel (1962) reviewed reports on 113 twin couples and found 95 per cent concordant homosexuality in the monozygotic twins and only 5 per cent concordance among the dizygotic twins. His evidence is in excellent agreement with Kallmann's data. Hutchinson (1959), in a broad theoretical discussion on the origins of paraphilia, has reemphasized Kallmann's work and viewed it, along with animal and other human studies, as leaving little theoretical doubt as to the existence of a genetic influence on sexual orientations. He suggests that gene-specific components may affect ". . . the rates of development of neuropsychological mechanisms involved in identification processes . . . ." Recently Melicow and Uson (1964), in an excellent review of human sex anomalies, have postulated that there may exist gene(s) in the sex chromosomes responsible for "identification and feel of maleness or femaleness." If the normal attachment of this gene is broken from the Y chromosome in males or the Y chromosome(s) in females, it may become transposed to another chromosome predisposing to abnormal sex role orientation. In a most recent work Kallmann (1963) reviews much of the more recent genetic data and still maintains that genetics are crucially involved in psychosexual development, the mechanism, however, still to be determined. |
|
|
Gedda (1963) has reevaluated Kallman's data statistically and claims to have computed that they show a minimum genetic contribution of 30 per cent in the etiology of homosexuality. |
|
|
Hampson and Hampson (1961) and J. L. Hampson (1964) have considered Kallmann's work, but regard his evidence as being outweighed by two types of studies. The first of these is exemplified by the work of Pare (1956). who, using the Barr technique for sex chromatin analysis, investigated male homosexuals for presence of the female chromatin body in the cell nucleus. Pare found no difference between the sex chromatin of the homosexuals and control males. This is a weak counter argument to the genetic one, since a negative finding of this sort is not necessarily evidence against the importance of genetic differences. With present techniques, chromosomal studies can do little more than indicate the presence of an extra sex chromosome or deficiency of an entire sex chromosome, or a gross translocation. They cannot reveal a single deviant gene or even several. The second type of study is more weighty, and consists of psychiatric and psychological reports which claim psychosexual disorders to be the result of social learning. These are exemplified by the reports of a multitude of workers who describe family situations and experiences which supposedly provide a basis for homosexuality. Green and Money (1961a, b) also attribute sexual deviation to certain kinds of experiences. In discussing the possible etiology of effeminacy in 5 prepubertal boys they cursorily review and dismiss the possible influence of a genetic involvement and prefer instead to consider only improper imprint experiences at an early age, despite the fact that common imprint experiences could not be proved. Gildea and Robins (1963) commented on the comparative findings of Kallmann and of Money in the following terms: |
|
|
His [Kallmann's] findings of perfect concordance for homosexuality in identical twins would indicate that homosexuality is in some way genetically determined and that the process of rearing cannot override the genetic constitution. Kallmann's findings do not fit very well with Money's emphasis on the crucial importance of rearing in determining sexual direction. |
|
|
Dobzhansky (1962) too comments on social experience, genetics, and human sexuality, thus: "Let us suppose that all persons who exhibit deviant sexual behavior as adults are found to have had certain kinds of experience in their childhood, particularly in their relation to their parents. But does it follow that such experiences induce sexual deviations in everybody who had them in his biography? Or does this happen only to the carriers of certain genetic endowments, which are apparently not rare in human populations?" Undoubtedly we are dealing with an interaction of genetics and experience; the relative contribution of each, however, may vary with the particular behavior pattern and individual concerned. Effeminacy and other sexual deviations are not altogether rare in our society. Without contending that inheritance is the whole cause, we may well consider it influential and predisposing. A quotation from Mead is again applicable: "It seems safe to assume that any behavior which can be institutionalized in a culture and regarded as a recurrent possible human choice has some hereditary base" (Mead, 1961). This hereditary base is what, especially for other animals, is usually termed an innate and instinctual framework within which the individual develops, meets, and contends with his environment. Most individuals, falling within the range of normal sexual behavior for our society, may be considered as following a path of genetic least resistance - that of their "innate instincts." In analyzing human behavior, trying to separate genetics and experience may be like trying to separate hydrogen and oxygen in their importance with respect to the properties of water. Those misassigned individuals who go against the predisposition inherent in their genetic make-up overcome it only to the extent their genetic flexibility will allow. Schultz (1963), in reviewing psychosexual problems associated with intersexuality and transvestism, stated in regard to the origin of psychosexual orientation that: |
|
|
The ultimate basis of hereditary endowment is influenced peristatically from the earliest time onward by the action of hormones which again are perhaps accessible not only to broadly somatic but even neuropsychiatric influences. Thus the love-life is fitted biologically to the dynamic totality of the constitutional make-up, and this unitaryformation corresponds in psychic range to the not less dynamic personality development. Even this rests on primary assumptions of a genotypic and certainly mainly primitive kind, of which the development, now thoroughly analogous to the development of constitution, is determined by an immense number of conditions. |
|
|
In the development of any behavior pattern we must consider not only the stimuli serving to mold the pattern but the nature of the receptive medium which is to be molded. Many humans, sibs in particular, can be subjected to extremely diverse experiences and still develop similar personalities; conversely, humans may be subjected to like experiences and develop quite differently. The individual's constitutional capacity to respond to these environmental stimuli exerts its influence, too. It is the genetic heritage of an individual which predisposes him to react in a particular manner so that the learning of a gender role can occur. Even if one thinks in terms of imprinting and innate releasing mechanisms (IRM) (Money, Hampson, and Hampson, 1957; Money, 1960a), these concepts must be recognized as implying that the individual possesses a genetic heritage. The IRM does not appear de novo but is species-specific and, as its name says, innate. Hormonal Organization of Sexual Behavior. To support the theory of psychosexual neutrality at birth we have been presented with no instance of a normal individual appearing as an unequivocal male and being reared successfully as a .female, or vice versa. Wherever the etiology of the hermaphroditism was determined the particular patients had been subjected, during their prenatal or neonatal existence, to a genetic or hormonal imbalance, or both (Hampson and Hampson, 1961). This imbalance may provide the requisite basis for what seems to be psychosexual neutrality. Schultz considers that in normal individuals there are in the two sexes differential psychosexual tendencies which determine the degree of male or female characteristics each individual will manifest. Hermaphrodites, however, he considers to possess an "undifferentiated constitution;" since they display a sexual erotic state low in vigor, and occasionally bordering on the eunuchoid, yet with nothing specific in incidence or character (Schultz, 1963). Benjamin (1964) reports that for transsexual individuals, "A certain significance endocrinologically may be attached to the fact that in 28 out of these 91 patients, a more or less distinct immaturity and hypogonadism to the point of eunuchoidism was found." |
|
|
["Transsexualism -a striking disturbance of gender role and gender orientation. It is a disorder of the harmony and uniformity of the psychosexual personality" (Benjamin, 1964).] |
|
|
It has long been recognized that various hormones are potent regulators of human behavior. Earlier cultures had a knowledge of the effects of castration, and much recent evidence is available to confirm the role that androgens serve in sexual drive and activity in men. Occasional studies to the contrary, the bulk of evidence indicates that testicular androgens activate libido, and that castration or other androgen loss due to some pathological condition reduces libido. Reviews (Lipschütz, 1924; Tauber, 1940; Beach, 1948a, b; Kinsey, Pomeroy, and Martin, 1948) and more recent case studies (Brewer, 1959) can readily document this conclusion. Brewer (1959), in evaluating the effect of castration in 157 cases, says: |
|
|
. . . in all cases without exception the amount of sexual activity has been altered. It has been reduced or abolished, irrespective of the direction or the form of sexual urge-heterosexuality, homosexuality, fetichism, zoophilic actions, masturbation, exhibitionism, or fetichistic actions . . . [emphasis original. |
|
|
Females too are influenced by their endocrines. The early work of Foss (1937), Shore, Papanicolaou, and Stimmel (1938), Loeser (1940), Geist (1941), Salmon and Grist (1943), Kupperman and Studdiford (1953), and the recent work from the Sloan-Kettering Institute is probably quite definitive in this respect (Waxenberg, Drellich, and Sutherland, 1959; Schon and Sutherland, 1960; Sopchak and Sutherland, 1960; Waxenberg, Finkbeiner, Drellich, and Sutherland, 1960: Waxenberg, 1963). These studies show the significant role androgens usually play in contrast to the "female" hormones, the estrogens and progesterone, in maintaining and stimulating erotic inclination, sexual desire, and behavior in females. In males the origin of the androgen is primarily from the testes, in the female from the adrenal and the ovary itself. The androgenic influences are acknowledged by Money (1961a). However, he sees them as supporting the neutrality theory, essentially on the following line of reasoning. Sexual desire and erotic functioning of both men and women are dependent upon a similar group of substances -androgens. Since men react to androgens with male sexual behavior and females react to androgens with female sexual behavior, the hormones are nondirectional, only activational, and the direction comes from another source. "The direction or content of erotic inclination in the human species is not controlled by the sex hormones. Hormonally speaking, the sex drive is neither male nor female but undifferentiated - an urge for the warmth and sensation of close body contact and genital proximity" (Money, 1961a). The direction taken by the drive is assumed to be learned or imprinted purely from the sex of rearing. Hampson and Hampson (1961) have stated that among patients whose sex hormones and secondary sexual body development contradicted the sex of rearing only 16 per cent were unable to adjust unambivalently to the assigned sexual role. Thus they reason that sex hormones do not act as causal agents in the establishment of an individual's gender role and psychosexual orientation. Perloff (1949, 1963), Kinsey and his associates (1948, 1953), and others also suggest that in the adult human individual endocrines are not the sole instigator or sole director (or both) of sexual behavior. Indeed, this position is not under contention. But to say that something is not the cause or sole director of a particular effect does not mean that it has little or no influence. Admittedly, hormones are probably not the single causal agent that induces gender role orientation, but their influence is undeniable and strong. Money (1961e) has said, "The sex hormones, it appears, have no direct effect on the direction or content of erotic inclination in the human species. These are assumed to be experientially determined." But the importance of the hormones in this regard was implicitly admitted, since hormonal therapy was recommended for correction of psychosexual and physical doubt (Hampson and Hampson, 1961). George W. Corner in his classic book The Hormones in Human Reproduction (1942) has said: |
|
|
Human [sexual] behavior involves all sorts of mental processes not subject to experimental control. We may be sure, however, that the hormones have an important part in the matter, directly or indirectly, and that without them there could be no human mating. |
|
|
These hormones, however, can affect behavior only to an extent that is inherent and previously organized within the soma. Boss (1943), in considering the activating properties of hormones in regard to a somatic base of sexual behavior, stressed that genetic factors are strongly involved, not only in determining the specific pattern of the taxonomic unit, but also of the sex. She considered this to be demonstrated by the widely if not generally valid rule that the same amounts of hormone do not produce identical reactions in the two sexes of a given species. Young (1961) has expressed a similar idea as follows: "We feel that the responses that the members of different sexes give to hormonal stimulation are predetermined; in this sense a specific rather than a non-specific relationship exists." Harris (1964) most recently presented the idea in this manner: "This (sex specific differential behavior response to hormones) reflects, in all probability, some anatomical or biochemical difference in the central nervous system of the two sexes." A most critical point to be elaborated here is that although endocrines may be primarily activational in the adult, in the fetus or neonate they must be considered directional. Phoenix, Goy, Gerall, and Young (1959) have clearly demonstrated that female guinea pigs are behaviorally as well as somatically masculinized by an androgen, testosterone propionate, prenatally injected into the mother. Postnatal behavior, more than the genitalia, was shown to be liable to alteration by the androgen, as even somatically unaffected females were behaviorally masculinized. Genetic females, if potentiated by androgens, manifested a suppression of the capacity to display lordosis following estrogen and progesterone treatment, and male-like mounting behavior was displayed by many of these animals even when lordosis could not be elicited (Phoenix, Goy, Gerall, and Young, 1959). Similar studies with the rhesus monkey afforded comparable results. that is, prenatally administered androgens will alter the normal female behavior patterns and genital structures to those of the male (Young, Goy, and Phoenix, 1964). These data may be taken as evidence of the organizing ability of prenatally acting androgens on the neural tissues mediating sexual behavior. Human females, in utero, also have been shown to be structurally modified by androgens (Wilkins, Jones, Holman, and Stempfel, 1958; Grumbach, Ducharme, and Moloshok, 1959; Grumbach and Ducharme, 1960; Diamond and Young, 1963). Although it has yet to be adequately demonstrated that these embryonic individuals were behaviorally affected, the possibility is within reason and not without zoological precedent. A possible mechanism for genes and hormones to interact and provide direction has recently been suggested. In contrast to the well-known evidences that genes can control hormonal activities, Karlson (1963) proposes and cites experimental evidence to show that steroid hormones are capable of altering and controlling gene activity. If such an alteration were to occur in cells within the (behavior mediating) nervous system we might expect an influence on subsequent behavior, certainly if the change were to occur during a critical prenatal period. Just as the presence of androgen has been demonstrated to be crucial, the prenatal absence of androgens is also crucial for the organization of mammalian tissues. |
|
| Although it has not yet been definitely shown that the morphogenic substance elaborated from the fetal testis is an androgen similar in character to adult testoids, most inferential evidence points in this direction (Bums, 1961). | |
|
Jost (1958) and Burns (1961), in excellent reviews, have cited the experimental evidence indicating that sexual differentiation of the normal male and female is dependent upon the presence of testicular substances. If testicular substances are present the differentiation is masculine; if absent, the differentiation is feminine. Males castrated prior to sexual differentiation develop structurally as females; in them there is an absence of external male genitalia and maintenance of derivatives of the Müllerian duct, e.g., the fallopian tubes, uterus, etc. Comparable human evidence is available from different types of hermaphroditic individuals; those with gonadal (testicular) aplasia (Grumbach, Van Wyk, and Wilkins, 1955; Grumbach and Barr, 1958; Jones and Scott, 1958: Wilkins, 1960; Overzier, 1963; Armstrong, 1964) and testicular feminization (Morris, 1953; Grumbach and Barr, 1958; Jones and Scott. 1958; Marshall and Harder, 1958: Barrio, 1962: Kendall and Loewenberg, 1962; Morris and Mahesh, 1963: Overzier, 1963; Armstrong, 1964) in particular. These genetic males are phenotypically female. Hampson. Hampson, and Money (1955) have claimed that since these individuals who possess a male chromatin pattern, can assume a gender role as normal females it means that the gender role is assumed independently of the genetic sex. It must be recognized that these genetic anomalies deprive the developing fetus of proper gonadal substances which in the human species, as demonstrably in other animals, may potentiate and organize the nervous system for masculine behavior. The deprivation of typical hormones or the presence of heterotypical ones may simultaneously potentiate and organize the nervous system for female behavior. Most significant are the findings which bridge the hormonal induction phenomenon discussed above and the resulting demonstrable sex-specific differences in the nervous system, in the hypothalamus in particular. Harris and Jacobsohn (1950, 1951), Martinez and Bittner (1956), Harris (1964), and Gorski and Wagner (1965) have shown that early in development the hypothalamus is differentiated into a male or female type. Their studies emphasize this in regard to the hypothalamo-hypophyseal axis in particular. Pituitaries transplanted beneath the hypothalamus from males or females into hypophysectomized castrated males will not cycle, whereas hypophysectomized females bearing either male or female pituitary grafts show complete and normal estrus cycles. Later Barraclough (1961), Barraclough and Gorski (1961, 1962), Harris and Levine (1962), Whalen and Nadler (1963), and Gorski and Barraclough (1963) demonstrated how this sexual differentiation of the hypothalamus may be achieved relatively simply with a single injection of steroid in a neonatal animal whose nervous system is still undifferentiated sexually. These experiments reflect on the organization of neural tissues implied in the work of Phoenix, Goy, Gerall, and Young (1959) and most probably occur in nature by means of the induction of gonadal substances. The implications are clear. The well-known and documented instances of hormonal regulation of sexual behavior below the human level are seen to be brought under neural mediation. With a broader outlook we see that genetic forces induce gonadal development, and gonadal development is normally followed by the elaboration of fetal or neonate gonadal substances responsible for the sexual differentiation of the nervous system. The neuroendocrine relations of the hypothalamus are the most obvious entities affected but it may be surmised that other, purely neural, aspects are also differentiated or potentiated at this time. Since most sexual anomalies are under genic control (Grumbach and Barr, 1958; Jones and Scott, 1958; Overzier, 1963; Armstrong, 1964; Beatty, 1964; Brunner-Lorand, 1964) it may be inferred that it is via such indirect and subtle means, as through hormonal action, that genes may be expected to influence organization of the nervous system and to mold sexual behavior. Thus rearing and assignment based on phenotype are not all that is involved in analyzing the etiology of sexual behavior in hermaphrodites. Those individuals with a male chromatin pattern who successfully assume a female gender role do so with the absence of crucial (hormonal) potentiating factors that may, in a normal male, establish a male constitution and a predisposition for the assumption of a male gender role. In the only behavioral studies of their kind to date, Grady and Phoenix (1963), and Feder and Whalen (1965), working with William C. Young, have completed studies which show that male sexual behavior is influenced by the loss of the neonatal gonad. They castrated male rats prior to sexual differentiation and observed that when reaching the normal age of maturity these rats fail to show masculine behavior and can readily be induced to show female behavior. Phoenix, Goy, Gerall, and Young (1959) aptly summarize these sorts of data: |
|
|
For the neural tissues mediating behavior, corresponding relationships [to the development of the genital tracts] seem to exist. The embryonic and fetal periods are periods of organization or "differentiation" in the direction of masculinization or feminization. Adulthood, when gonadal hormones are being secreted, is a period of activation; neural tissues are the target organs and mating behavior is brought to expression. Like the genital tracts, the neural tissues mediating mating behavior respond to androgens or to estrogens depending on the sex of the individual, but again the specificity is not complete (Antliff and Young, 1956; Young, 1961). |
|
|
They go on to discuss the question of inherent neural bisexuality as follows: |
|
|
Like Dantchakoff (1938a, b, c), Raynaud (1938), and many others (Beach, 1945a; Steinach, 1913, 1916; Lipschütz, 1924; Beach, 1942, 1945b), the existence of a bisexuality is assumed. We suggest, however, that in the adult this bisexuality is unequal in the neural tissues as it is in the case of the genital tissues. The capacity exists for giving behavioral responses of the opposite sex, but it is variable and, to most mammals that have been studied and in many lower vertebrates as well, it is elicited only with difficulty (Young, 1961). |
|
|
Therefore, when we consider prenatal as well as postnatal existence, hormones may be regarded as directional as well as activational; and at birth the individual may be considered to have been neurally predisposed by genetic and hormonal means toward one sex. Since this predisposition is demonstrated so vividly in animals, including anthropoids, we may logically assume that it persists in the human after birth, although manifestations of it may be suppressed or modified. In a recent review article Young, Goy, and Phoenix (1964) expressed such confidence about this point that in relation to the theory of Hampson and Money they stated: "In view of what we have learned an endocrinological basis which is consistent with the concept of psychologic bisexuality exists for the interpretation of most if not all of the cases they report." The Nervous System. In discussing behavior, we find it inevitable to consider in some detail its medium - the nervous system. The point at issue here is whether the nervous system is just a neutral responsive system intermediate between stimulus and response, or is a system with a built-in diasthetic bias. This consideration is crucial, since it is on the nervous system that potentiation and organization must exert their influence and it is on the nervous system that learning (to be discussed below) exerts its effect. Previously, genetic and hormonal factors were seen to be capable of organizing the nervous system to direct future sexual behavior. More direct, immediate, and independent influences of the nervous system may be understood from clinical studies and from experiments investigating behavior by means of ablation, lesion placement, direct stimulation, direct application of hormones, electrical self-stimulation, or electrical recording. These various methods reveal different functional levels of activity and the sites of so-called "sex-centers" - loci for the integration of the component activities of a particular sex behavior pattern. Significantly, these studies often reveal inherent differences of the nervous system dependent upon gender. Were a "neutral" nervous system to exist, identical effects would be expected from individuals of both sexes. The majority of direct studies do not set out primarily to investigate response differences between the sexes. These differences are in fact taken for granted, and sex-specific response patterns such as lordosis and estrus behavior are used as dependent variables. The functional role of a specific locus or structure is thus usually analyzed only in relation to the anticipated patterns applicable to the sex of the animal investigated. But although male-female comparisons are not usually made, direct studies are of value in illustrating that loci within the nervous system possess inherent properties related to sexual disposition, behavior, and manifestations of gender role. The most significant differentiation of the hypothalamus has been mentioned above in discussing the role of the endocrines in sexual differentiation. The hypothalamus has for quite some time been implicated both indirectly and directly in influencing sexual behavior patterns. Its indirect functioning via the hypophysis has been discussed by Harris (1955, 1956, 1960, 1964), Green (1956), Greer (1957), Sawyer (1960, 1962), Szentágothai, Flerkó Mess, and Halász (1962), Nalbandov (1963), Everett (1964), and others. The significance of this functional pattern was discussed in an earlier section (p. 161). Direct functioning without hypophyseal involvement has been suggested by the experiments of Brookhart and Dey (1941), Clegg, Santolucito. Smith, and Ganong (1958), Sawyer and Robison (1956), Robison and Sawyer (1957), Soulairac (1959), Soulairac and Soulairac (1956), Phoenix (1961), and Goy and Phoenix (1963). These studies inferring direct hypothalamic involvement indicate a sex-specific functioning of certain loci. Crucial for our argument are instances where similar loci were investigated in both males and females. For example, Goy and Phoenix (1963) have demonstrated that midventral hypothalamic lesions in female guinea pigs result in an alteration of the normal sexual behavior ranging from a complete loss of the capacity to display estrus to permanent estrus, depending upon the precise locus of the lesion. In male guinea pigs (Phoenix, 1961) similar lesions also produced behavioral alterations. The type of alteration is significant. Lordosis was lost in the female, and mounting was lost in the male. Many reviews (Beach, 1951; Goldstein, 1957; Sawyer, 1960, 19112) detail the relative dependence of the male and independence of the female on the cortical mass in relation to the competent expression of sexual behavior. Complete removal of the cortex will not prevent mating responses in the female rat, cat, rabbit, or guinea pig, whereas destruction or removal of 75 per cent of the male cortex completely abolishes the male mating pattern. In female rats which spontaneously exhibit male-like mounting, decortication stops this behavior whereas the usual female behavior components are unimpaired (Beach, 1943). While it may be argued that most of the data are from nonhumans, Ford and Beach (1951) have suggested that the human male and female are also differentially dependent upon cortical mass relative to their expression of sexuality. These, then, are indices of sexually differentiated nervous tissues. The classical work of Klüver and Bucy in 1939 and by Klüver (1952) afterwards showed that sexually mature male macaques respond to bilateral removal of the temporal lobe by hypersexual activity: heterosexual, homosexual, and autosexual in direction. The female does not show this. Significantly, however, Klüver (1952) remarked, ". . . an intensification of sexual responses may occur even in a pseudohermaphrodite or in a female from which the uterus and both ovaries have been removed prior to extirpating the temporal lobes." Later in his review Klüver (1952) added, "There is no doubt that age, sex, species and many other factors influence the picture of behavior alterations produced by a bilateral temporal lobectomy." Schreiner and Kling (1955, 1954, 1956) and Green, Clemente, and De Groot (1957) have similarly demonstrated that lesions in the piriform cortex of the male cat induced hypersexual changes, whereas hypersexuality was not usually observed in female cats with similar lesions. Analogous functioning of the human cerebrum may be inferred from clinical reports of the Klüver-Bucy syndrome (Sawa, Ueki, Arita, and Harada, 1954; Terzian and Dalle Ore, 1955) and other disorders of the temporal lobe (Epstein, 1960, 1961). Interestingly, these reports show sexual deviation manifested only in men. Lansdell has reported a sex difference in verbal ability (1961) and in design preference (1962), which reacts differentially to temporal lobe surgery. After surgery to the dominant lobe, women maintain their previous "artistic judgment" while men lose theirs- "The effects of the operations suggest that some physiological mechanisms underlying artistic judgment and verbal ability may overlap in the female brain, but are in opposite hemispheres in the male" (Lansdell, 1962). On a more subtle level, perceptual differences between males and females can be considered. In children, the work mentioned earlier with Rorschach tests (Ames, Learned, Métraux, and Walker, 1952) and block building (Ames and Learned, 1954) is suggestive of the presence within the nervous system of perceptual and cognitive processes which are early manifested sex differences. On a purely neurophysiological basis Lipsitt and Levy (1959) have shown that as early as the first three days of life females show a lower threshold to electric shock stimulation than do males. Adult women usually have greater olfactory acuity than men (Schneider and Wolf. 1955) and this has been seen to vary with hormone levels (Schneider, Costiloe, Howard, and Wolf, 1958). Kinsey et al. (1953) reported that males, more often than females, are erotically oriented to visual cues. Females may be more susceptible to tactile stimuli. Sexual distinctions such as these cannot help but influence various aspects of an individual's life, including the way he or she views and reacts to the world, and hence gender role. It may be argued, as Money (1963x) does, thatdifferences such as perceptual responses are results of learning rather than inherent in the nervous system and causal. However, Money presents no model to explain how or why perceptual thresholds to stimuli which are obscure and novel, such as those used by Lansdell, may be differentially altered and learned, particularly where the patient is unaware of a desirable direction for the change. Because of the nature of the tests and the early ages at which they were administered, it would also be difficult to explain how the Rorschach and block-building tests were biased by learning. It may be concluded that the nervous systems of males and females are differentially reactive to the environment. And there is good reason to believe that these differences, so definite in humans as well as in other animals, are present and influential at birth and afterwards. Imprinting. Hampson and Hampson (1961) contend that: |
|
|
The premise that behavior is based primarily on instincts is gradually disappearing from scientific writing and the traditional concept of instinct is undergoing revision and modification. In its place has emerged the view that early experience importantly structures subsequent behavior. This is not to say, lest misunderstanding arise, that the animal organism - human or subhuman - is merely a blank slate to be written upon by the capricious finger of life experiences. Quite the contrary, for there are now many studies in the literature dealing with genetic constitution and the inheritance of basic capacities affecting later learning, temperament and personality. |
|
|
This theme is developed so that the constitutional factors are considered insignificant when compared with ontogenetic factors. Difficulty in incongruous sex orientation or modification of sex is thought of in ontogenetic terms of crucial timing and imprinting. To support their view, Beach and Jaynes (1954) were cited by Hampson and Hampson (1961) to show how early experiences may modify behavior. Emphasis was placed on the last of three possibilities given by Beach and Jaynes, namely, that which postulated a "critical period" in ontogeny during which certain types of behavior are irrevocably structured for the remainder of the organism's life. However, once stated, the possibility was accepted as an established fact. Hampson and Hampson allow, it is true, for species variation, but now consider this variation to be minor. |
|
|
Neither the Hampsons nor Money give criteria to indicate when they think species differences (between humans and other animals) are negligible. They are grossly inconsistent in this respect. In relation to basic sexual tendencies, the human is assumed to be quite distant even from other mammals. But in relation to imprinting, seen definitely only in birds, men and birds are assumed to be quite close. |
|
|
Hampson and Hampson have not recognized that Beach and Jaynes mentioned, in the same paragraph, when referring to critical periods and imprinting in humans and birds: "It is tempting to suggest the existence of similarities in underlying mechanisms, but this would be unwarranted on the basis of present knowledge" (Beach and Jaynes, 1954). Schiller (1957) in Instinctive Behavior, isolated imprinting as predominantly avian and contrary to mammalian behavior. Fletcher (1957), in his book Instinct In Man, although discussing human sexuality at length, did not even consider the possibility that imprinting might be involved in the behavior of humans. Nevertheless, the "psychosexual neutrality-at-birth theory" assumes the human species, in regard to the establishment of gender role, to be analogously imprinted (Money, Hampson, and Hampson, 1957; Money, 1960a, 1961b, d; Hampson and Hampson, 1961). Money has even gone so far as to say, "It is quite likely that aberrations of sexual inclination and behavior, the so-called perversions, are errors of imprinting;" and that "the phenomenon of falling in love can be analyzed as an imprinting phenomenon" (Money, 1960a). Since no experimental studies on the imprinting of human sexuality per se are available, animal experiments may provide further insight into such a possibility. Young (1961), in reviewing isolation experiments with the guinea pig, stated, "The data as a whole indicate that the emergence of sexual behavior patterns in the male guinea pig is not restricted to an early critical period comparable with that described in the literature dealing with imprinting (Lorenz, 1937)." Rats, too, have been shown to be relatively free from the need of sexual imprinting (Beach, 1958), and so have mice (King, 1957). Since lower mammalian species are free of an imprinting requirement for competent sexual behavior, it seems that it would be difficult to defend the proposition that humans, a behaviorally more flexible species, are subject to such an imprinting stereotype. Admittedly, although many species and individual variations exist (Scott, 1962), "critical periods" do exist for mammals in regard to reproductive phenomena. But while reproduction may be affected, assumption of a sexual role may not be. A normal rhesus monkey raised in isolation may not react sexually sufficiently to mate, but a male still follows male behavior patterns (albeit deviantly); and a female, female patterns. Harlow (1961) has mentioned that many types of sexually dimorphic behavior patterns such as play, aggression, and receptivity, which are as much a part of the proper gender role as coitus, arise spontaneously in isolated male and female monkeys. He said, "It is unreasonable to account for these sex differences as learned, culturally ordered patterns of behavior because there is no opportunity for acquiring a cultural heritage, let alone a sexually differentiated one, from an inanimate cloth surrogate" (Harlow, 1961). This conclusion is in contradistinction to the response of the female rhesus which, as mentioned earlier, when potentiated by prenatal androgen injections, behaves not as a deviant female but as a male. There seems to be little doubt that normal humans would manifest sexually dimorphic behavior patterns even if raised in isolation. It is interesting here to consider the following point. If imprinting were involved to any great extent and the gender role independent of a more significant sexual instinct, then a neutrality theory would have to contend with the fact that for the first three years of life babies are almost exclusively with their mothers (or some other female), and yet male children are usually not imprinted as females. This fact must be considered in the light of the observations that imprinting, as we see it in birds, occurs within the brief space of a few hours and irrespective of sex. Nothing really comparable to imprinting has been demonstrated to occur in human sexuality. Learning and Reinforcement |
|
|
The concept of reinforcement is important and basic to any learning theory. A reinforcement is any change in the organism or in the environment which has the ability to increase the probability of occurrence of some particular behavior performed just prior to the presentation of the reinforcement (Thorndike, 1913: Hull, 1952). Thus, orgasm may be reinforcing to an animal and increase the probability of some action required to obtain the orgasm. |
|
|
Much of the psychosexual "neutrality at birth" theory depends upon the great part that learning plays in the instillation of masculine and feminine gender roles (Hampson and Hampson, 1961: Money, 1960x, 1961e). Sears, Maccoby, and Levin (1957), Rabban (1950), and others are cited by Hampson and Hampson (1961) as indicating that culture and learning shape subsequent behavior. Ehrmann (1963) has recently provided an excellent review of various studies investigating the social determinants of human sexual behavior. These studies indeed indicate that gender personality is influenced by society and thus, in part, is learned. But Ehrmann wisely started his review by emphasizing that in looking for the social determinants of sexual behavior one must take into account biological considerations and not simply learning processes. Benjamin (1964) in a very recent study of transsexual individuals could find no evidence that childhood conditioning is involved in the etiology of the transsexualism in 47 out of 87 patients. Of the remaining 40 patients conditioning was of "doubtful" influence in 24 individuals and definitely influential in only 16 cases. As a function of learning, assumption of a gender role presumably should follow the characteristics of a normal learning curve. But, when viewed properly, modifiability of sex behavior by learning must be seen also as a disproof of the imprinting theory, for the same modifications of behavior cannot be caused both by learning and by imprinting in the same species. The basic incompatibilities are that learning requires reinforcement while imprinting does not, and that imprinting is fixed and irreversible while learning is not. But even to grant that assumption of a gender role by an individual in our society may be molded by learning in no way proves that it is the sole or dominant determiner of the gender role. Money, Hampson, and Hampson would have us believe so. Hampson and Hampson (1961) contend: |
|
|
In the human psychological sexuality is not differentiated when the child is born. Rather, psychological sex becomes differentiated during the course of the many experiences of growing up, including those experiences dictated by his or her own bodily equipment. Thus, in the place of the theory of an innate, constitutional psychological bisexuality such as that proposed by Freud-a concept already questioned on theoretical grounds by Rado (1940), among others - we must substitute a concept of psychologic sexual neutrality in humans at birth. Such psychosexual neutrality permits the development and perpetuation of divers patterns of psychosexual orientation and functioning in accordance with the life experiences each individual, may encounter and transact. Rado (1940) indeed questions the concept of bisexuality, but nowhere does he assume neutrality at birth. Instead, he claims that there is at birth a differentiated unisexuality. He states, "Under normal developmental conditions, as differentiation proceeds and one type of reproductive action system grows to completion, the original bipotentiality ceases to have any real significance .... The standard developmental pattern of our species provides for each individual only one reproductive action system. The two inherent potentialities of the zygote are thereby mutually exclusive." |
|
|
It is not obvious, when we examine the best possible evidence, how the criteria of learning can be logically applied to the hermaphroditic evidence. Yet to do just that is the core of their philosophy, which asserts that if gender roles are modified by learning or are established by imprinting, then sexual behavior patterns are not prenatally mediated. Here is a subtle non sequitur. All evinced learning, be it operant or classical, is by definition a modification of behavior (Verplanck, 1957). The evidence presented in this paper indicates that the learning of a gender role is a culturally fostered ontogenetic phenomenon of development superimposed on a prenatally determined pattern and mechanism of sexual behavior. As Tinbergen (1951) has stated. "There is a close relationship between innate equipment and learning processes, in that learning is often predetermined by the innate constitution. Many animals inherit predispositions to learn special things, and these dispositions to learn therefore belong to the innate equipment." This predisposition may be considered also in terms of differential endocrine sensitivity. The nervous systems of males and females subjected to genetic and hormonal factors may present an altered learning capability that sets limits to or extends the range of sexual behavior possible to the individual. Hutchinson (1959) has proposed that the individual's genetic endowment operates to mold behavior by affecting "the rates and extent of development of the neuropsychological mechanisms underlying the identification process and other aspects of object relationships in infancy." An extensive search of the literature reveals no case where a male or female without some sort of biological abnormality, e.g. chromosomal, hormonal, or gonadal, accepted an imposed gender role opposite to that of his or her phenotype. If such an individual is available he has not been referred to by proponents of a "neutrality at birth" theory. It may be assumed that such an individual will be hard to find, since this anomalous behavior would be outside normal limits. Human hermaphrodites, it would seem reasonable to assume, are somehow altered in their neurophysiological capacity towards or away from certain features of sexual behavior. But since their behavior still falls within the broad scope of normal behavior their actions usually go unchallenged. It might also be pertinent here to state that, in humans, behavior can be affected and that for females passivity may serve as "normal" sexual behavior and thus seriously hamper any rating system of "femaleness." An individual's personal conviction as to his or her sex, even if bolstered by suitable genitalia, may lack the essential reinforcing properties of orgasm. For females this normally does not seem to be crucial (Wallin and Clark, 1963), but for males orgasm may be a necessary reinforcement of the sex role (Ford and Beach, 1951). This greater dependence of males upon orgasm as reinforcement may be significant in a more general way, in that males are more capable of sexual learning and conditioning, whereas females seem to show a relative absence of such processes (Ford and Beach, 1951). Parenthetically, we may mention the existence, of differences in genital response, only recently investigated. These could hardly have been learned. Masters and Johnson (1968) have shown that practically all stages of the orgasmic process differ between males and females. Clitoral erection is comparatively slow during initial excitement; then the clitoris retracts and essentially maintains a "buried" position until the orgasmic phase has passed. Only after the cessation of vaginal orgasmic contractions does the clitoris reappear (Masters and Johnson, 1963). So although we may speak of the penis and clitoris as homologous structures, their functioning is not parallel. In truth, instinct is modified by learning as much as learning is modified by instinct (Tinbergen, 1951; Verplanck, 1955; Deese, 1958). The evidence may well indicate that while among all species sex roles and the learning of them are prenatally organized, such roles, particularly among humans, may be greatly modified though never fully negated. It is natural to conclude that both biological and cultural factors interact and meld to determine masculine and feminine gender roles (Dengrove, 1961). To say that learning predominates in sex role assumption, when based upon evidence from hermaphrodites, is unwarranted. All this evidence may show is that hermaphrodites are quite flexible in their assumption of a gender role. The surprise or abnormality would be if humans, whom we consider to be among the highest evolved forms of life, could not greatly modify instinctive behavior. To jump the gap by extending the breadth of this modifiability and to say that because of it man is completely divested of his evolutionary heritage to instinctive mediation is specious. Tinbergen (1951), in discussing the instincts and causation underlying human behavior, said, "Mating behavior in man, not in the form of the accomplishment of the consummatory act, but in the preparatory, appetitive stage of 'lovemaking' proves, when studied ethologically, to be basically dependent on sex hormones and on external stimuli and it is on these agents that our rational powers exact a regulating influence." Beach (1949) has offered a basis from which human sexual behavior may be studied when he said, "To interpret the sexual behavior of men and women in any society it is necessary first to recognize the nature of any fundamental mammalian pattern and then to discover the ways in which some of its parts have been suppressed or modified as a result of social pressures brought to bear upon the individual" (emphasis added). The proper interpretation of human sexual behavior must not stumble on a debate of nature versus nurture. Undoubtedly both are significantly involved! SUMMARYThe evidence and arguments presented show that, primarily owing to prenatal genic and hormonal influences, human beings are definitely predisposed at birth to a male or female gender orientation. Sexual behavior of an individual, and thus gender role, are not neutral and without initial direction at birth. Nevertheless sexual predisposition is only a potentiality setting limits to a pattern that is greatly modifiable by ontogenetic experiences. Life experiences most likely act to differentiate and direct a flexible sexual disposition and to mold the prenatal organization until an environmentally (socially and culturally) acceptable gender role is formulated and established. It is also indicated, on the one hand, that the argument in favor of an imprinting type of assumption of gender role is spurious for the human being and that, on the other hand, learning theory cannot account for all observed sexual behavior. That imprinting does not account for assumption of the gender role is shown by the failure of the phenomena to meet the criteria for imprinting and by the inability of an imprinting theory to handle various facets of human behavior. That learning cannot account for all sexual behavior was shown by reviewing the original evidence, and by presenting clinical, comparative cultural, and animal studies. Instead, human sexual behavior is seen to be a composite result of prenatal and postnatal factors. The evidence derived from human hermaphroditism, instead of constituting a separate group of data from which a state of psychosexual neutrality at birth is to be inferred, can perfectly well fit into the classical and phylogenetically consistent theory of sexual predisposition at birth. This conclusion is best seen not when the evidence is atomized, and gender role is compared with each atom of influence, but when all the evidence is considered in toto. Considering normal psychosexual development, the two theories may be represented as in Fig. 1. In Fig. 1, A. a psychosexually neutral origin provides a narrow basis from which all the manifestations of sexuality are supposedly developed. In comparison, Fig. 1, B depicts the comparatively broad basis of a sexually predisposed individual. The former theory postulates specific imprints or learning experiences which would enlarge and direct sexual development. The latter need only postulate cultural restrictions and learning to narrow and concentrate sexual direction and orientation. As the figure indicates, there are various "critical stages" which affect the range of subsequent sexual capability. A theory of psychosexual neutrality sees these stages as levels which remove the individual further from the neutral midline. A theory of psychosexual predisposition views these stages as imposing limits and restrictions in the form of culturally and biologically acceptable sexual outlets within the total capability. To cross from the male to the female "path" in Fig. 1, A would mean to overcome a relatively broad gap (indications of an irreversible change): for malassigned individuals to cross over in Fig. 1, B would require only transcending a culturally imposed barrier to arrive on a different biologically "predisposed" path. All in all, the second model (B) seems best suited to the available anthropological and clinical data, which predicate the initial existence of a bisexuality that is subsequently narrowed. [Chall (1963) presents a scheme of levels which is an attempt to depict graphically the determinants of human sexual behavior.] 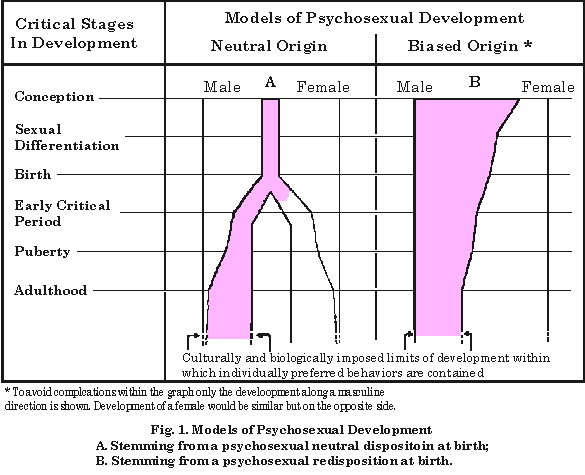
CONCLUSIONThe theory of inherent sexual predisposition and of a somatic basis for the patterning of sexual behavior is not original with me. Aside from mythological and religious beliefs of a similar nature, this hypothesis was advanced scientifically almost fifty years ago by Goodale (1918). It was scientifically revived by Ball (1937) and by Young, Dempsey, Myers, and Hagquist (1938) over twenty-five years ago. Recently Young (1961) has reviewed much of these data, but he did not consider the human species. Essentially, I have added an accumulation of evidence bringing the human species into line with others in respect to the existence of a somatic basis for sexuality. The clinical aspects of the work on which their theory is based, and the case management and therapy recommended by Hampson, Hampson, and Money, as well as by Wilkins (1964) and others, may, in the main, be applicable to our society and appropriate. This aspect of their work was not under discussion. However, doubt as to the validity of their theory may reasonably raise some question as to its proper extension to practice. H. F. Bettinger considers that a hermaphrodite should be given treatment, medical and sociological, in accordance with the sex of personal preference, and irrespective of the sex of rearing. He stated (Bettinger, 1950): |
|
|
In practice this formula has been usually applied to those patients who were brought up as girls but developed distinct male feelings during adolescence. They have been advised to change their legal sex and to undergo surgical operations aimed at approximating as closely as possible their anatomical structures to the male pattern. On the whole, their management has been quite successful, mainly for the reason that it was held that their 'true sex' was male, and that therefore, everything done for them to make them more like ordinary males was right and proper and deserved every encouragement. |
|
|
Other clinicians have expressed similar convictions (Armstrong, 1955: Norris and Keetel, 1962; Dewhurst and Gordon, 1963: Berg, Nixon, and MacMahon, 1963). Benjamin (1964) advises similarly for the treatment of transsexuals. Brown and Fryer (1957), clinicians at the Washington University School of Medicine, claim never to have seen a single male patient who did not eagerly anticipate and undergo the change from sexual malassignment to the correct status regardless of the degree of distress over the original error. Kraft and Bedford (1963) have even set forth a methodology for facilitating sex reassignment when describing the change of a five-year-old "boy" to a girl without psychopathological sequelae. Thus, from a practical standpoint alone, the theory and practice advocated by the Hampsons and by Money may be questioned. In a most recent and excellent review on the psychiatric aspects of intersexuality, Roth and Ball (1964) have stated: |
|
|
The anatomic equipment will limit or dictate what [treatment] can be undertaken but the patients social circumstances, sexual identification and personal wishes have to be allowed to decide the issue in the majority of cases as far as this is practicable. |
|
|
A competent and extensive clinical reappraisal is perhaps warranted. It is time, not as Hampson and Hampson (1961) have said, "to assume sexual neutrality and [that] the theory of bisexuality must be laid to rest when one considers the evidence;" but rather to admit that in sexuality, as in so many other areas, the human being is extremely flexible and his behavior is a composite of prenatal and postnatal influences with the postnatal factors superimposed on a definite inherent sexuality. With this approach we can advance toward an analysis of the mechanism of this composite formation and begin to analyze further the interplay of phylogeny and ontogeny in human sexual behavior. ACKNOWLEDGMENTSI am indebted to Dr. Robert W. Goy who originally suggested the writing of this paper, and to Drs. William C. Young, Charles H. Phoenix, and Arnold A. Gerall for enlightening discussion of the theories and problems involved in a presentation of this type. Naturally, they are in no way responsible for any shortcomings herein. This study was supported in part by the Committee for Research in Problems of Sex, National Academy of Sciences-National Research Council. and in part by Public Health Service Research Grant GM 10632 of the National Institutes of Health, Public Health Service. |
|